Bone marrow mononuclear cells for joint therapy: The role of macrophages in inflammation resolution and tissue repair
2021-07-30BrunoMenarimJamesMacLeodLindaDahlgren
Bruno C Menarim, James N MacLeod, Linda A Dahlgren
Bruno C Menarim, James N MacLeod, Gluck Equine Research Center, Department of Veterinary Science, College of Agriculture, Food and Environment, University of Kentucky, Lexington,KY 40546, United States
Linda A Dahlgren, Department of Large Animal Clinical Sciences, Virginia-Maryland College of Veterinary Medicine, Virginia Tech, Blacksburg, VA 24060, United States
Abstract Osteoarthritis (OA) is the most prevalent joint disease causing major disability and medical expenditures. Synovitis is a central feature of OA and is primarily driven by macrophages. Synovial macrophages not only drive inflammation but also its resolution, through a coordinated, simultaneous expression of pro- and anti-inflammatory mechanisms that are essential to counteract damage and recover homeostasis. Current OA therapies are largely based on anti-inflammatory principles and therefore block pro-inflammatory mechanisms such as prostaglandin E2 and Nuclear factor-kappa B signaling pathways. However, such mechanisms are also innately required for mounting a pro-resolving response,and their blockage often results in chronic low-grade inflammation. Following minor injury, macrophages shield the damaged area and drive tissue repair. If the damage is more extensive, macrophages incite inflammation recruiting more macrophages from the bone marrow to maximize tissue repair and ultimately resolve inflammation. However, sustained damage and inflammation often overwhelms pro-resolving mechanisms of synovial macrophages leading to the chronic inflammation and related tissue degeneration observed in OA. Recently,experimental and clinical studies have shown that joint injection with autologous bone marrow mononuclear cells replenishes inflamed joints with macrophage and hematopoietic progenitors, enhancing mechanisms of inflammation resolution,providing remarkable and long-lasting effects. Besides creating an ideal environment for resolution with high concentrations of interleukin-10 and anabolic growth factors, macrophage progenitors also have a direct role in tissue repair. Macrophages constitute a large part of the early granulation tissue, and further transdifferentiate from myeloid into a mesenchymal phenotype. These cells, characterized as fibrocytes, are essential for repairing osteochondral defects.Ongoing “omics” studies focused on identifying key drivers of macrophagemediated resolution of joint inflammation and those required for efficient osteochondral repair, have the potential to uncover ways for developing engineered macrophages or off-the-shelf pro-resolving therapies that can benefit patients suffering from many types of arthropaties, not only OA.
Key Words: Osteoarthritis; Arthropathy; Synovitis; Hematopoietic progenitor; Homeostasis; Cell therapy
INTRODUCTION
Osteoarthritis (OA) is the leading joint disease that affects people and domestic animals and causes physical disability and substantial medical costs[1-3]. The pathogenesis of OA is not fully understood; however, chronic synovial inflammation is crucial in the progression of OA and is frequently the sole driver of related degenerative changes[4,5]. The complexity of the inflammatory response in the synovium, ultimately causing degeneration of synovial tissues during OA, represents a critical therapeutic challenge. Commonly used non-steroidal anti-inflammatory drugs (NSAIDS) and intra-articular corticosteroids provide marked clinical improvement during earlier stages of OA. However, such outcome is a consequence of antagonizing cellular pathways that block not only the inflammatory reaction, but also cellular pathways fundamentally involved in efficient tissue repair and recovery of homeostasis (e.g., nuclear factor kappa-B signaling)[6,7]. Therefore, they can have detrimental effects on cartilage metabolism that go far beyond chondrocyte quiescence. Corticosteroids often inhibit the endogenous production of cytokines by resident macrophages [e.g., interleukin (IL)-10 and prostaglandin E2(PGE2)] that are required for optimal chondrocyte function and other joint cells favoring tissue repair and a return to synovial joint homeostasis[7-10].
One way to cease inflammation and its degenerative consequences, avoiding the negative side effects of anti-inflammatories is to support endogenous resolution of the inflammatory process. Importantly, inflammation resolution is a natural process that requires biological events that happen during the onset of acute inflammation. Hence,the blocking of acute inflammation using NSAIDS or corticosteroids can interfere with efficient inflammation resolution, often leading to chronic inflammation[11-13].Biologicals such as mesenchymal stem cells (MSC), autologous conditioned serum and platelet rich plasma offer pro-resolving advantages over anti-inflammatories[14-19],however, improvements observed from these therapies have short-lasting effects.Therefore, two major goals are imperative in developing more effective treatments of inflammation in OA: (1) Preserve cellular and molecular mechanisms involved in the physiology of joint tissues; and (2) Favor long-term resolution by supporting those components of the inflammatory response that efficiently clear the underlying triggers of inflammation as detailed below.
There is cumulative evidence that macrophages are the central drivers of the inflammation in the synovium of OA-affected joints[20-23], but also the central drivers of inflammation resolution[11,12,24-28]. This review focuses on the dual role of macrophages in the maintenance of joint homeostasis, alternatively inducing joint inflammation, and the emergent therapeutic use of macrophages from the bone marrow [within bone marrow mononuclear cells (BMNC)] to promote endogenous resolution of joint inflammation and recovery of homeostasis.
SYNOVIAL MACROPHAGES – FIRE FIGHTERS IN SYNOVIAL INFLAMMATION
Two cell types represent over 80% of the cellular composition of the synovial membrane: Synovial macrophages and synovial fibroblasts, historically known as type A and type B synoviocytes, respectively. Most cells in the synovium are synovial fibroblasts (type B, about 55%). Synovial macrophages are the second most predominant cell in normal synovium (type A, about 25%), and are observed in higher density in the sub-intima as compared to the intima[21,29,30]. Increases in numbers of both cell types can result from inflammation. During the early stages of OA, synovial macrophages are the predominant cell type in the synovial lining (about 50%);however, with disease progression fibroblasts tend to dominate (about 70%)[21,31]. In end-stage OA, synovial cells are replaced by fibrous tissue impairing the basic functions of the synovium[32].
Macrophages are the cornerstone of synovial inflammation. Although synovial fibroblasts and articular chondrocytes can intensify the inflammatory response in the joint, they cannot primarily incite it without macrophages[21,33,34]. As a matter of fact, macrophage depletion during experimental arthritis[35] or clinical cases of rheumatoid arthritis[36] confer clinical improvement and a marked decrease in the expression of OA biomarkers, underscoring the central role of macrophages in driving synovial inflammation[20,21,35-37]. Also, macrophage activation in the osteoarthritic synovium has been correlated to pain and disease activity and severity[38,39].Macrophages are, however, also essential for optimal chondrocyte function, inflammation resolution, and thus joint homeostasis[23,40]. In healthy joints, macrophages promote synovial homeostasis through phagocytosis (i.e., clearance of foreign bodies,debris, and apoptotic cell removal), secretion of synovial fluid growth factors,chemokines and cytokines, as well as other paracrine effects supporting chondrocyte function and required for the terminal differentiation of chondrocyte progenitors[21,23,40]. Synovial macrophages can also line up along the edges of the synovial lining,forming a tight junction-mediated defense system for the communication between the intra- and extra-articular environments[41]. Therefore, macrophage depletion impairs vital elements of joint integrity.
Following any kind of tissue damage, resident macrophages first respond by delimitating the injury, creating a shield under which they further drive tissue repair[42]. During such a response, macrophage activation results in an increased production of IL-1, IL-6, tumor necrosis factor (TNF)-α and several alarmins, triggering a reparative response to damage. This reaction is immediately followed by equivalent increases in expression of IL-10, insulin-like growth factor (IGF-1) and other anabolic and growth factors[43-46], leading to decreased production of these pro-inflammatory cytokines and providing an ideal scenario for tissue repair to progress[43]. If the extent of the injury overwhelms the capacity of macrophages to fully surround the lesion,macrophages “blow the whistle”, recruiting neutrophils readily available from the blood stream and setting the stage for a short-term, acute inflammatory response[42].This acute inflammatory response in turn induces macrophage recruitment from the bone marrow in addition to proliferation of resident macrophages, amplifying the macrophage-mediated process of tissue repair and inflammation resolution[11,12]. If none of these responses prove sufficient to repair damaged tissues and ultimately resolve inflammation, this cellular and cytokine feedback loop of pro- and anti-inflammatory mediators is sustained. Nonetheless, in situations of extensive or continuing tissue damage, the sustained inflammatory response decreases the capacity of macrophages to produce anti-inflammatory, or more correctly, pro-resolving molecules (e.g., IL-10 and IGF-1), impairing mechanisms of inflammation resolution and leading to chronic inflammation[45,47]. Ultimately, the perpetually increased production of cytokines and activation of catabolic enzymes causes further damage or aberrant remodeling of joint tissues[5,20,48].
In summary, when the homeostatic functions of synovial macrophages become overwhelmed, they upregulate inflammation to ultimately recruit more macrophages and respond to the increased demands for repair[21,41,42,49]. Efficient resolution of inflammation is required to interrupt the catabolic processes in OA and to re-establish a homeostatic synovial environment[48,50] (Figure 1). Understanding the facts outlining the phenotypical response of synovial macrophages in both the initiation and resolution of joint inflammation is paramount for developing pro-resolving joint therapies[11,20,21,40,50].
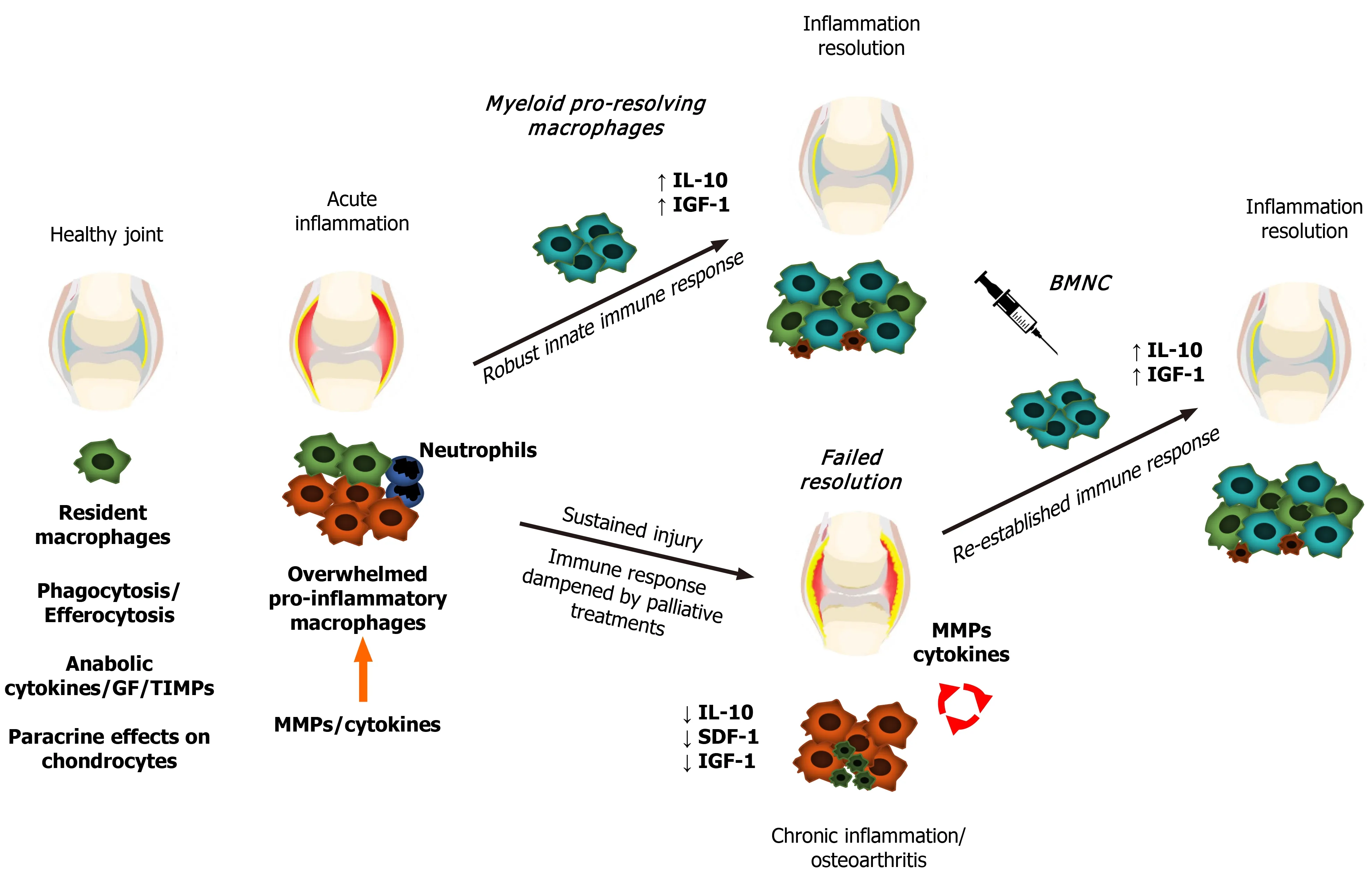
Figure 1 Schematic representation of macrophage status and responses during joint health and disease. Macrophages promote synovial health through phagocytosis, efferocytosis, secretion of synovial fluid cytokines and growth factors, and paracrine effects on chondrocyte metabolism. Following damage,synovial macrophages form a shield to the injured site to further drive tissue repair. If the extent of the injury or continuing trauma overwhelms the shielding and repairing capacity of macrophages, macrophages increase expression of cytokines, including interleukin (IL)-1, IL-6, and tumor necrosis factor -α in a phlogistic inflammatory response, recruiting first neutrophils, followed by myeloid macrophages. These macrophages have a strong pro-resolving capacity associated with high production of IL-10 and insulin-like growth factor 1, which then leads to inflammation resolution and tissue repair. If interfering factors impair the resolution of the inflammatory process, chronic low-grade inflammation persists, leading to aberrant remodeling of synovial tissues and joint degeneration. Joint injection with bone marrow mononuclear cells in chronically inflamed joints augments the macrophage-mediated mechanisms of joint homeostasis, resolving joint inflammation. GF:Growth factors; TIMPs: Tissue inhibitors of metalloproteinases; MMPs: Matrix metalloproteinases; IL-10: Interleukin 10; IGF-1: Insulin-like growth factor 1; SDF-1:Stromal-derived factor 1; BMNC: Bone marrow mononuclear cells.
Macrophage phenotypes in the synovium − beyond M1 and M2 polarization
Following challenge under definedin vitroconditions, macrophages activate into a spectrum of phenotypes, where the extremes are characterized by cells exhibiting classical pro-inflammatory (M1) or pro-resolving/healing (M2) responses[51].In vivo,macrophages respond to variable environmental stimuli, exhibiting a dynamic range of phenotypes that varies by tissue and health status[12,27,51-53]. In synovial joints,classically activated (M1-like) macrophages from the osteoarthritic synovium impedeex vivochondrogenesis of synovial progenitor cells, while alternatively activated (M2-like) macrophages are required for efficient chondrogenesis[40,54] and inflammation resolution, improving clinical and histological signs of joint disease[49,55].Collectively, these findings suggest that promoting the M2-like response in diseased joints may provide a therapeutic tool to favor inflammation resolution and tissue repair. Taken together, these observations suggest that harnessing the response of macrophages to enhance the ability of the joint to resolve inflammation can help in the treatment of OA and other joint diseases.
Until recently, specific data about macrophage phenotypes in OA were limited toin vitrostudies, end stage OA or experimental animal models[21,40,54,56,57]. Comparative analysis of phenotypical responses between macrophages from healthy and osteoarthritic joints included inferences from other types of arthritic conditions[58].OA related reports were limited to low amounts of macrophages in synovial fluid,which shed from the synovial membrane after hyperactivation or mechanical separation, and thus are unlikely to reflect the response within the synovium[59,60].
To better understand the roles of macrophagesin vivo,we recently assessed the patterns of macrophage phenotype activation in the synovium from healthy equine joints (n= 29) and compared to that of joints with naturally occurring OA (n= 26)[47].We evaluated the synovial expression of widely used M1-, M2-like and pan macrophages markers [cluster of differentiation (CD)14 (pan-marker), CD86 (M1-like),CD206, and IL-10 (M2-like)] and correlated these findings with synovial histology.Joints with moderate OA were selected to represent a disease stage where the cellular response in the synovium is high, largely attributable to synovial macrophages and also more likely to respond favorably to treatment[61]. We then correlated the findings to cytokine/chemokine profiles in the synovial fluid. Macrophage phenotypes in the synovium were not as clearly defined as they arein vitro. All macrophage markers were expressed with minimal differences between OA and normal joints. However, in OA joints these markers increased proportionate to synovial inflammation, especially CD86. These findings were associated with hyperplasia of the synovial lining, which reflects increased macrophage recruitment and activation in response to injury[47,62].
Among 12 cytokines assayed in that study[47], concentrations of stromal cellderived factor (SDF)-1 and IL-10 were lower in synovial fluid from osteoarthritic joints,while macrophage chemoattractant protein (MCP)-1 was higher. Upon inflammatory stimuli, synovial fibroblasts release high concentrations of MCP-1. Elevated MCP-1 in synovial fluid from osteoarthritic joints, combined with hyperplasia of the synovial intima characterizes a fibroblast-mediated recruitment of macrophages to the synovium in response to injury[63]. On the other hand, SDF-1 is an essential chemokine in the recruitment of macrophages during inflammation resolution[47,64-66]. Macrophages are the main source of IL-10[7,67], an essential cytokine for cartilage homeostasis and tissue repair[8,9,68,69]. In the face of increased macrophage recruitment, the observation of decreased concentrations of SDF-1 and IL-10 suggests that mechanisms of macrophage-mediated inflammation resolution may be compromised or overwhelmed during OA.
In conclusion, macrophage phenotypes observedin vivoare more diverse and complex than the clear classifications described throughin vitrostudies[27,39,56,57].In vivo, synovial macrophages are neither M1 nor M2, but a hybrid phenotype that overall exerts a homeostatic response to injury by driving inflammation to counteract tissue aggressors and trigger tissue repair[27,41,42,46,55,70-74]. Cytokines and cell surface markers frequently used for assessing phenotype identity (M1- or M2-like)invitroare actually building blocks of a multifaceted immune response and must be prudently analyzed together and in consideration to the dynamic stages of the inflammatory reaction, including its resolution[11,73,75]. Why endogenous recruitment from myeloid macrophages to the joint often fails in recovering synovial homeostasis is not yet known; however, epigenetic mechanisms derived from chronic synovial inflammation seem to affect the performance of local macrophages[76-78]. Developing approaches to recover the homeostatic mechanisms from healthy macrophages may yield improved therapeutic options to prevent or resolve joint inflammation in OA joints and re-establish joint health.
BONE MARROW MONONUCLEAR CELLS AS A THERAPEUTIC SOURCE OF HEALTHY MACROPHAGES
BMNC have been investigated in the fields of tissue repair and treatment of chronic inflammation in several non-arthritic conditions over the last 20 years. BMNC therapy in the management of chronic airway inflammation in murine models and equine clinical cases resulted in clinical improvements comparable to treatment with corticosteroids and associated with marked increases in IL-10 production[7,24]. The management of patients with cerebral palsy using intra-thecal injections of BMNC substantially improved gross motor function and muscle spasticity and tone[79-81].BMNC have also improved functional recovery from acute liver failure and related survival in mouse models[82-84]. Finally, and impressively, BMNC made possible the successful transplantation of pancreatic islet in both murine and primate models[85].There have also been some studies reported with skeletal tissues. BMNC treatment shortened the inflammatory phase of healing and improved tissue quality in tendons and ligaments[86-89], as well as in osteochondral defects[90-93]. Taken together, the beneficial effects described suggest that autologous BMNC therapy may be an attractive option for chronically inflamed tissues and those with limited regenerative capacity.
Due to its autologous nature, BMNC therapy eliminates the risk of graft-versus-host disease. Because BMNC are readily isolated from bone marrow and do not require culture expansion, BMNC does not present the adverse reactions associated to cell culture neither are subject to the regulatory restrictions that apply to MSC, and are FDA approved for use in people[94]. BMNC isolation from bone marrow aspirates using gradient centrifugation is rapid and can be completed with minimal manipulation. Cells are readily available for immediate administration using few equipment[90,95,96]. The resulting BMNC cell isolate is composed primarily of mononuclear cells(> 90%). Although most of these cells are macrophage- and monocyte-committed progenitors (< 50%), there are also hematopoietic stem cells (about 25%), lymphoid cells (about 10%), fibroblastic reticular cells (about 10%), and a very small portion of MSC (0.00001%)[24,97,98]. It is the macrophage progenitors in BMNC that are primarily responsible for resolving inflammation. Other hematopoietic and MSC contribute, but to a lesser extent[24,84]. Moreover, the MSC therapeutic effects are largely associated with activating a homeostatic response in macrophages, and thus BMNC represents a more direct, targeted approach[10,99]. Given the central role of synovial macrophages in joint health and inflammation, it only makes sense to capitalize on the macrophage-mediated effects of BMNC to re-establish mechanisms of joint homeostasis to develop a targeted OA therapy. Even though the clinical use of purified macrophages has been suggested as a great opportunity to improve patient outcomes[100], BMNC retains a large component of hematopoietic and progenitor cells (about 25%) that serves as a reservoir for the self-renewal of large amounts of macrophages “consumed” during inflammation and its resolution[98].
Bone marrow mononuclear cells therapy for joint disease: Scientific and clinical evidence
During the last decade, several reports have demonstrated successful use of BMNC in the treatment of chronic synovitis and joint disease[25,46,55,101-104]. Early studies suggesting BMNC could be an effective therapy for joint disease evaluated its effect on“tennis elbow” (elbow lateral epicondylitis), and found a significant improvement using a standardized patient-rated evaluation. Unfortunately, the experimental design did not include an assessment of control groups or a comparative therapy[101]. More recently, Goncarset al[102], evaluated the clinical efficacy of a single joint injection with BMNC to improve pain and other symptoms of moderate knee OA [Kellgren-Lawrence (KL) stage II–III] in a cohort of 28 patients that were followed for 12 mo[104]. The clinical assessments were based on the Knee osteoarthritis outcome score(KOOS) and the Knee society score (KSS) at baseline, 3 mo, 6 mo, and 12 mo after injection, and compared to a patient group (n= 28) treated with 3 consecutive weekly injections of sodium hyaluronate[104]. Significant improvement (P< 0.05) was observed in the BMNC-treated group for all scores at all time points, with special superiority in the KOOS pain subscale for BMNC over the hyaluronic acid-treated group at 6 and 12 mo.
In a second study, the same authors focused on correlating the changes in knee OA symptoms (KL grade II and III) with magnetic resonance imaging (MRI) changes following treatment with a single BMNC injection in a slightly larger cohort (n= 34)[102]. Clinical outcomes were again analyzed using the KOOS and KSS systems at baseline, 3 mo, 6 mo, and 12 mo after injection. Results from MRI were measured using the Whole Organ Magnetic Resonance Imaging Score at baseline and 6-7 mo post injection. Remarkably, 97% of joints treated with BMNC exhibited clinical improvement that was still perceived 12 mo post-treatment in at least 65% of the patients, which still exhibited a difference of more than 10 KOOS points from baseline.Improvements were significant (P< 0.05) for both clinical and imaging scoring systems. Of note, MRI findings revealed that most patients had improvements in synovitis (63%) and bone marrow edema (57 %), but most remarkably, improvements in cartilage were also observed in 38% of patients (Figure 2). No imaging data were available for hyaluronan-treated patients from the previous study. The authors concluded from the second study that a single dose of BMNC (45.56 × 106± 34.94 × 106BMNC containing 1.04 × 106± 1.61 × 106CD34+ hematopoietic stem cells) reduced clinical signs of moderate knee OA, and in some cases, decreased degenerative changes of the joint tissues. Of note, a positive correlation between cell quantity (total BMNC or CD34+ cells) and the improvement in OA symptoms was not observed[102].
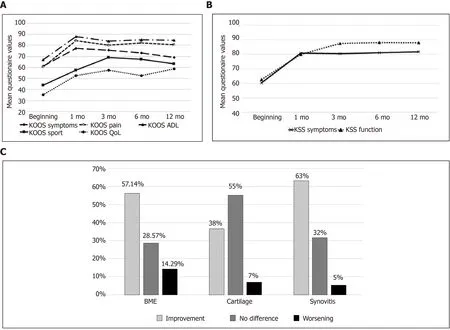
Figure 2 Clinical (Knee injury and osteoarthritis outcome score and Knee society score, n = 34) and imaging (magnetic resonance imaging, n = 30) outcomes from patients with knee Osteoarthritis treated with bone marrow mononuclear cells over 12 mo post-treatment[102]. A and B: Significant improvements (P < 0.05) were observed for Knee injury and osteoarthritis outcome score’ (A) and Knee Society Score (B) scores during the 12-mo follow-up period and most were sustained over time; C: Changes in bone marrow edema, cartilage and synovitis detected by magnetic resonance imaging 6 mo after treatment with bone marrow mononuclear cells. Citation: Goncars V, Kalnberzs K, Jakobsons E, Enģele I, Briede I, Blums K, Erglis K, Erglis M, Patetko L,Muiznieks I, Erglis A. Treatment of Knee Osteoarthritis with Bone Marrow–Derived Mononuclear Cell Injection: 12-Month Follow-up. Cartilage 2019; 10 (1): 26-35.Copyright © The Author(s) 2018. Published by SAGE Publications. KOOS: Knee injury and osteoarthritis outcome Score; KSS: Knee society score; QoL: Quality of life; ADL: Activities of daily living; BME: Bone marrow edema.
Our group recently completed a couple of studies aimed at understanding how normal and inflamed synovial joints respond to injection with BMNC[55], and how BMNC respond to an inflamed environment to produce a positive effect[46]. Special attention was paid to the macrophage component in BMNC. First, using a wellestablished equine model, we induced synovitis in both radiocarpal joints of 6 horses using lipopolysaccharide. Following 8 h, at the peak of the acute inflammatory response, one inflamed radiocarpal and one normal tarsocrural joint received BMNC injection (20 x 106viable BMNC, about 79% viability). Saline was injected in the contralateral joints. Synovial fluid was collected at 1 d, 4 d, and 6 d post treatment for cytology, flow cytometry for expression of macrophage markers, and cytokine quantification. Six days post BMNC therapy, following euthanasia, joints were assessed for gross pathology, and the synovium was harvested for histology and immunohistochemistry targeting markers of macrophage activation. At 4 d following treatment with BMNC, inflamed joints exhibited 24% more macrophages with 10% higher counts of IL10+ cells than saline-treated controls. Overall, BMNC-treated joints showed gross and analytical improvements in synovial fluid and synovial membrane, with increasing pro-resolving macrophages and IL-10 concentrations in synovial fluid compared to saline-treated controls. Inflamed joints treated with BMNC were histologically equivalent to healthy joints, whereas saline-treated controls remained abnormal[55] (Figure 3).
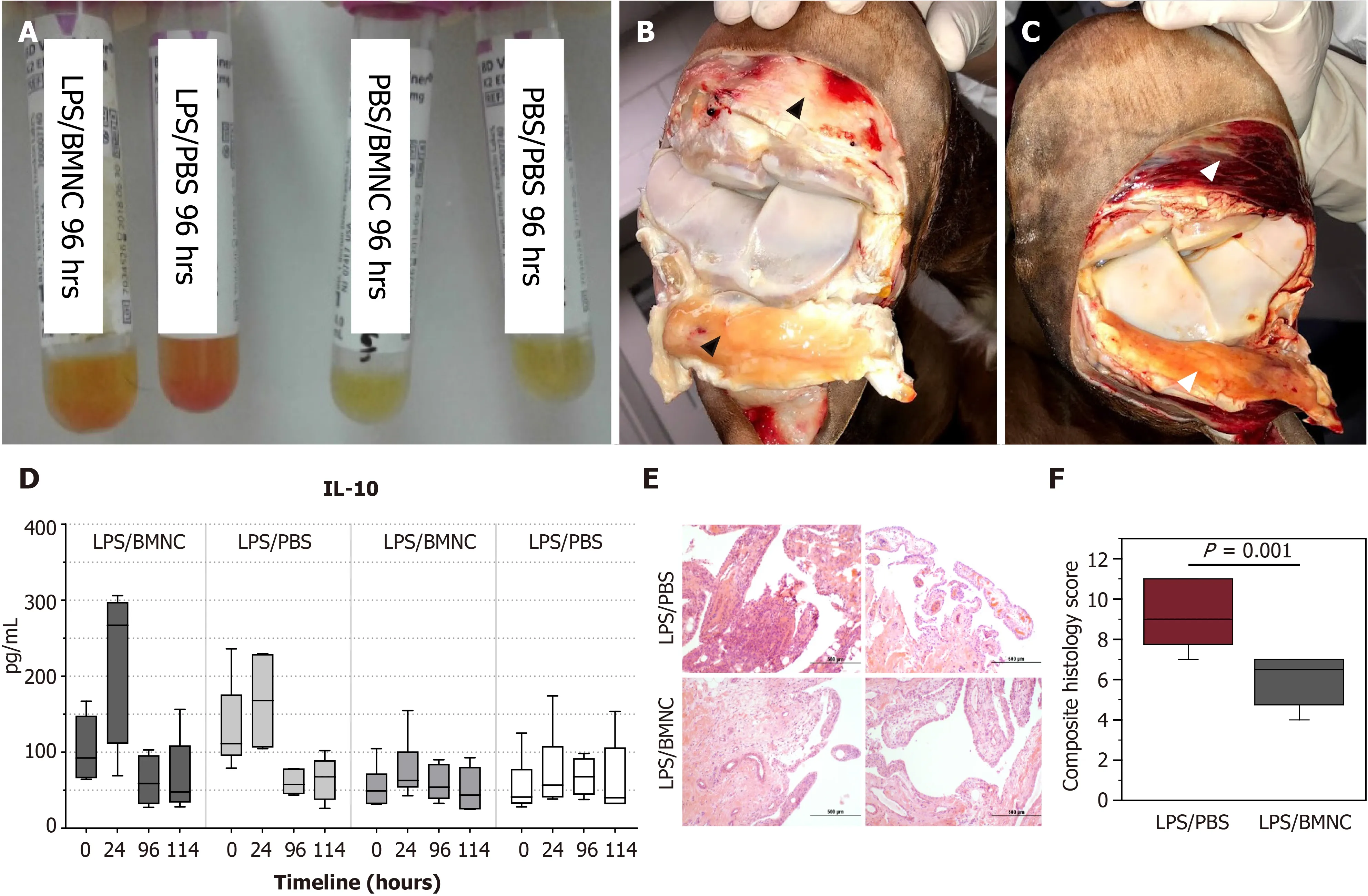
Figure 3 Gross and analytical findings from normal and inflamed equine joints treated with bone marrow mononuclear cells[55]. A:Improvements in synovial fluid associated with decreased cellularity and red cell contamination at 96 h compared to Phosphate buffered saline (PBS)-treated controls; B: Improvements in the synovium were reflected by decreased intra-and peri- and articular synovial hemorrhage and edema, in bone marrow mononuclear cells (BMNC)-(black arrowheads) compared to; C: PBS-treated joints (white arrowheads); D: There was a marked increase in synovial fluid interleukin 10 concentrations at 24 h in inflamed joints, which was much higher in BMNC-treated joints as compared with PBS-treated joints; E and F: BMNC-treated joints exhibited lower scores for all histological aspects of inflammation, although these were only significant (P < 0.05) for vascularity and the composite score(F); Citation: Menarim BC, Gillis KH, Oliver A, Mason C, Ngo Y, Were SR, Barrett SH, Luo X, Byron CR, Dahlgren. Autologous bone marrow mononuclear cells modulate joint homeostasis in an equine in vivo model of synovitis. FASEB J 33, 14337–14353. Copyright © The Author(s) 2019. Published by Wiley Online Library in cooperation with the Federation of American Societies for Experimental Biology. LPS: Lipopolysaccharide; PBS: Phosphate buffered saline; BMNC: Bone marrow mononuclear cells.
To understand how BMNC induced such therapeutic effects, we studied thein vitroresponse of BMNC to culture in normal (SF) and inflamed autologous synovial fluid(ISF) using cells and synovial fluid from the same horses from the just describedin vivostudy[55]. Equine BMNC cultured in SF or ISF (n= 8 horses) developed into macrophage-rich cultures exhibiting phenotypes similar to macrophages native to synovial fluid from healthy joints. BMNC confluence (cell proliferation) was ultimately higher in ISF (about 100%) than SF (about 25%). BMNC cultured in SF or ISF were neither M1- nor M2-like but exhibited a range of hybrid phenotypes with a proresolving response, characterized over time by decreasing secretion of IL-1β, gradually increasing secretion of IL-10 and IGF-1, and increasing counts of IL-10+macrophages.These changes were sustained over ten days and were more evident in ISF, suggesting that macrophage-mediated mechanisms of homeostasis were conserved over time and were likely favored by the gradual increase in cell proliferation. A combined assessment of data from ourin vivoandin vitrostudies suggests that joint injection with BMNC can increase the number of synovial macrophages and magnify the macrophage- and IL-10-associated mechanisms of joint homeostasis impaired during the progression of OA[45,46]. Moreover, BMNC therapy preserved, bothin vivoandin vitro, the production of cytokines required for tissue repair (PGE2, IL-10 and IGF-1), the same ones generally impaired by corticosteroids[46,55].
To more fully assess the effects of BMNC therapy in the treatment of naturally occurring equine OA, we conducted a small clinical study evaluating the response of joints (metacarpophalangeal and carpi) with moderate OA to BMNC therapy in 19 adult horses[25]. In the absence of a KL-equivalent scoring system for equine OA,scores were defined by consensus between 6 experienced clinicians. At baseline, horses were subjected to a clinical and musculoskeletal exam, including objective gait analysis at the trot on days 0, 7, and 21 post-treatment with either saline, BMNC (20 x 106viable cells) or a commonly used dose of triamcinolone (4 mg/joint). After treatment, all horses were rested in stalls for 1 wk, followed by 7 d of stall rest with hand walking,and then a resumption of normal activities in the third week. Lameness decrease was significant (P< 0.05) and consistent only in the BMNC-treated group (between 7 d and 21 d) and associated with increasing counts of macrophages in the synovial fluid.Combined, findings from all of the above-mentioned studies support that BMNC therapy can boost the homeostatic mechanisms of synovial macrophages critical for inflammation resolution.
BMNC as a therapy for osteochondral repair
The use of bone marrow components in the treatment of musculoskeletal conditions dates back from over a century ago and is the foundation for bone marrow grafting as today’s most used techniques for the treatment of non-union fractures[105-107] and osteochondral healing[90,91,108,109]. Newer methods based on the original grafting studies aim at providing means for cells from the subchondral bone marrow to reach the osteochondral defect through microfracture or subchondral bone drilling[109-113].For therapeutic purposes, removal of most red blood cells and bone spicules from bone-marrow aspirates through centrifugation was a further refinement of the technique because these components were identified as causing severe joint inflammation[114] and heterotopic bone formation[87]. The product resulting from centrifugation of bone marrow aspirate, rich in myeloid leukocytes and other hematopoietic and mesenchymal progenitor cells, is called bone marrow aspirate concentrate(BMAC). The leukocytes included in BMAC, granulocytes and monocytes, not only play crucial roles in inflammation, but also contribute to cellular mechanisms driving tissue repair[11,115-117]. However, the association of neutrophils with chronic inflammation and delayed repair lead clinicians and scientists to avoid their inclusion in biological joint therapies[115]. After removing bone spicules, red blood cells and granulocytes, the mononuclear fraction from the bone marrow (BMNC) is what is left and retains the osteochondral repair properties of bone marrow[90,94]. This is true even when the subchondral bone is not drilled or picked to allow subchondral bone marrow to reach the defect, suggesting that the cells responsible for osteochondral repair are within that BMNC fraction of the bone marrow aspirate[92,118].
The extent of which myeloid macrophage progenitors directly participate in osteochondral repair remains to be defined, but findings from other tissues suggest it goes beyond phagocytic and paracrine activities. Following damage and the subsequent neutrophil-driven acute inflammation that triggers wound healing, the injury site is further populated by mononuclear cells[117,119,120]. These cells display specific monocyte/macrophage markers and are responsible for removing neutrophils by efferocytosis and for the early production of collagen and enzymes required for remodeling the developing granulation tissue, which is largely composed by macrophages[49,121,122]. Following this initial phase, these same cells expressing macrophage markers trans differentiate into cells displaying a different set of markers typically seen in fibroblasts. Cells displaying this ability to trans differentiate are called fibrocytes, are abundant in the bone marrow, and have a central, intrinsic role in tissue development and repair[121-123]. Ongoing research may unveil the transcriptional switches driving these cells to trans differentiate from myeloid to mesenchymal phenotypes, uncovering targets to work on the optimization of cell-based therapies for cartilage regeneration.
Key quality control steps for harvesting and processing BMNC
Isolation of BMNC from bone marrow aspirates is traditionally done by gradient centrifugation. However, the volume of bone marrow aspirate reported for processing of BMNC varies from 5 to 60 mL and are extrapolated from data optimized for the isolation of MSC[124,125]. Lack of BMNC-specific data may adversely impact the cellular composition of the final BMNC product. To identify an optimal volume of bone marrow aspirate for BMNC isolation, Correa-Letelieret al[126], compared the concentration of BMNC in fractioned bone marrow aspirates in 16 horses. Bone marrow aspirates were obtained in 5 consecutive 5 mL fractions, and the mononuclear cell content of each fraction was quantified using differential cell counts. The total number (median ± QD) of mononuclear cells in the first three fractions (first 15 mL)contained 93.2% of the BMNC in the total 25 mL aspirated, with an average cell viability of 97.5 ± 1.2% (Figure 4). To the authors’ knowledge, similar comparative data for bone marrow aspiration in people is not available. The authors emphasize that, in horses, it is necessary to obtain at least the first 15 mL of bone marrow aspirate from the sternum when the sample is intended for BMNC isolation. Importantly, exceeding the total volume of 25 mL increases the peripheral blood content in the bone marrow aspirate, which has a poor concentration of the pro-resolving cells that are typically found in the bone marrow niche[125,126]. The result is a relative dilution of the desired BMNC with peripheral cells. One useful parameter in assessing if the bone marrow aspirate volume is excessive and diluted by peripheral blood is the platelet count, given that the concentration of platelets in the bone marrow is very low[96]. A detailed protocol for harvesting equine bone marrow aspirates and processing BMNC with minimal resources have been described elsewhere[96]. While only 20-40 x 106BMNC are used for joint injection, a bone marrow aspirate of 25 mL yields 150-200 x 106BMNC and the exceeding cells can be cryopreserved for future treatments without compromising BMNC function[127].
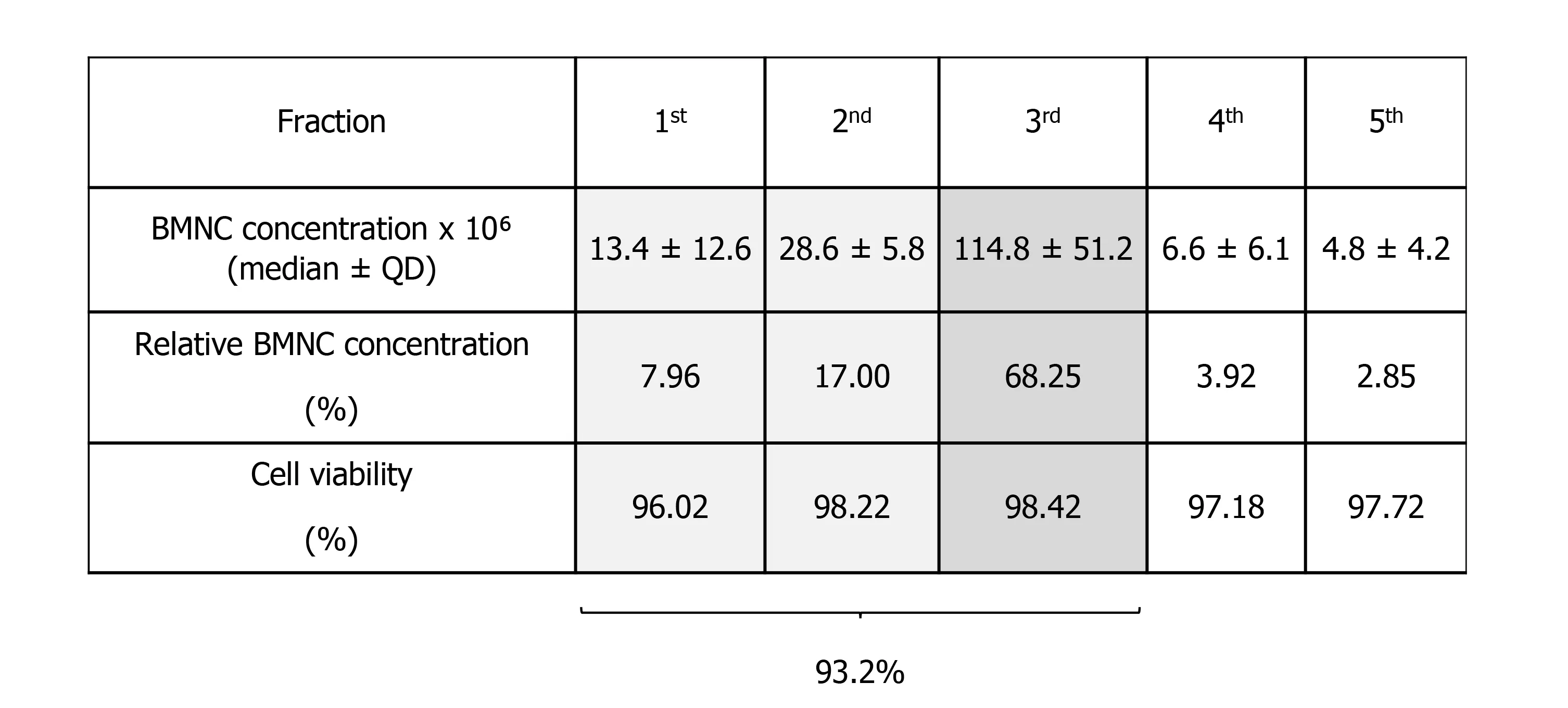
Figure 4 Mononuclear cell content in five consecutive bone marrow aspirates (5 mL each). The first three fractions represent 93.2% of the total 25 mL (the five fractions together) emphasizing the need to obtain at least the first 15 mL of bone marrow aspirate. BMNC: Bone marrow mononuclear cells.
CONCLUSION
Therapeutic strategies to promote the endogenous resolution of synovial inflammation in joint disease has the potential to avoid the negative side-effects of anti-inflammatory joint therapies. Despite substantial research efforts, there are remarkable gaps in knowledge concerning mechanisms for natural recovery from inflammation following injury and the re-establishment of joint homeostasis. Both inflammation and its resolution, are processes largely mediated by macrophages. Therefore, understanding macrophage behavior and function through the dynamic process of inflammation and its resolution is paramount for advancing this field and optimizing emergent cellbased therapies. Moreover, molecules and events involved in the inflammatory process of OA are frequently and inadvertently seen as causative of inflammation and thus labeled as having detrimental effects. However, the same mediators involved in initiating inflammation are also, either by themselves or by inducing the synthesis of other mediators and signaling mechanisms, necessary to effectively drive endogenous resolution of the inflammatory process. We do not yet understand why endogenous recruitment of myeloid macrophages to the joint during inflammation is often inadequate to recover homeostasis. However, BMNC therapy is a proven and easily accessible alternative treatment for OA that enhances the innate homeostatic mechanisms of synovial macrophages, providing long-lasting and pro-resolving effects. Ongoing “omics” studies may reveal key drivers of macrophage-mediated resolution of joint inflammation. Identifying new targets to explore pro-resolving mechanisms of therapeutic potential is a logical approach for developing engineered macrophages and perhaps off-the-shelf pro-resolving therapies to benefit patients suffering from many different types of arthropathies, not only OA.
杂志排行

World Journal of Stem Cells的其它文章
- Epigenetic modulators for brain cancer stem cells: Implications for anticancer treatment
- Mechanisms involved in selecting and maintaining neuroblastoma cancer stem cell populations, and perspectives for therapeutic targeting
- Roles of mitochondrial unfolded protein response in mammalian stem cells
- Stem cell therapies in tendon-bone healing
- Exosomal microRNAs from mesenchymal stem/stromal cells:Biology and applications in neuroprotection
- Immunotherapy against programmed death-1/programmed death ligand 1 in hepatocellular carcinoma: Importance of molecular variations, cellular heterogeneity, and cancer stem cells