红河干热河谷不同植物光合作用及抗逆生理特性1)
2021-07-30窦沛彤刘方炎高成杰李昆
窦沛彤 刘方炎 高成杰 李昆
(中国林业科学研究院资源昆虫研究所,昆明,650224)
干热河谷主要存在于我国西南横断山脉的金沙江、澜沧江、红河和怒江的中上游地区,在这些重要河谷地区形成了干湿季分明、降水少,高蒸发以及热量高的干热河谷气候,表现出“既热又干”基本环境特点[1-2],由于比较恶劣的自然环境,导致该地区干旱频发。
在干热河谷生长的植物经常遭受干旱胁迫及光胁迫,这影响了该地区植物正常的生长及发育。近些年,学者对干热河谷的研究主要集中在植物群落特征、多样性及树种的抗旱和光合特性等方面[3-12],其中孙永玉等[10]通过盆栽控水法对干热河谷柠檬桉苗期的抗旱生理进行了研究,郑元等[11]通过比较干热河谷不同种源的牛角瓜苗期生理特性,筛选出了高水平光合生产与水分利用潜力的牛角瓜种质资源,赵广等[12]通过对金沙江干热河谷区辣木(Morin-gaoleifera)、银合欢(Leucaenaleucocephala)、和罗望子(Tamarindusindica)的光合生理特性进行分析,认为在干热河谷植被恢复中需谨慎选择银合欢。目前,对红河干热河谷区代表性植物在自然条件下对干旱和强光胁迫的生理适应性研究还需要进一步完善,红河干热河谷旱季的环境条件是持续的干旱和高温,这是对植物生存的最大考验,也是植被恢复时的难点。本研究以该区域乡土植物天干果(BuchananialatifoliaRoxh. Hort. Beng)、山黄麻(Trematomentosa(Roxburgh) H. Hara)、虾子花(Woodfordiafruticosa(L.) Kurz)及车桑子(Dodonaeaviscosa(L.) Jacq.)为试验材料,探讨干热河谷地区不同植物的光合特性和抗逆生理特性等特征,以期揭示不同植物光合生理、抗氧化酶活性及渗透调节物质对干旱环境的响应,探索不同植物对干热河谷环境的适应性,为研究红河干热河谷地区的植被恢复、抗旱树种的选择提供科学参考。
1 研究地概况
研究区域位于云南省红河县境内(23°19′38″~23°24′55″N,102°15′1″~102°25′36″E),地处云南省东南部。研究区域处于低纬度亚热带高原型湿润季风气候区,每年5—10月为雨季,降水时空地域分布极不均匀。地带性土壤以红壤和燥红土为主,干旱瘠薄,水肥条件极差,在自然状态下以草丛为主,杂以灌木,稀少乔木,称为稀树(灌)草丛,地形地貌特殊,焚风效应强烈,形成了典型的干热气候特征[13-15],年平均干燥度1.7,属于半干旱偏湿亚类型[16]。
2 材料与方法
2.1 光响应曲线拟合
在2019年4月底,选择晴朗天气上午09:00—11:00,采用LI-6400XT便携式光合测定仪测定植物的净光合速率(Pn)。人工光源为LI-6400红蓝光源,设置光合有效辐射梯度为1 800、1 500、1 200、1 000、800、500、200、120、100、80、50、20、0 μmol·m-2·s-1,启动光响应自动测定程序。每种参试植物选择健康、长势均一的树3棵,每棵树选择3片叶,结果取平均值。
用SPSS 23.0对测定的净光合速率和光合有效辐射数据采用非直角双曲线模型进行拟合[17-18],得出相应的光合响应参数,拟合方程为:
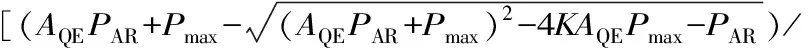
2K]-Rday。
式中:Pn为净光合速率;AQE为表观量子效率;PAR为光合有效辐射;Pmax为最大净光合速率;K为光响应曲线曲角;Rday为暗呼吸速率。在光合有效辐射≤200 μmol·m-2·s-1的低光强度下,净光合速率与光强存在正比线性关系,该直线方程与X轴的交点为光补偿点(LCP),当Pn为Pmax时所得的PAR为光饱和点(LSP)[19]。
2.2 气体交换测量
在2019年4月底,采用LI-6400XT便携式光合测定仪测定参试植物的净光合速率(Pn)、蒸腾速率(Tr)、光合有效辐射(PAR)、空气相对湿度、气温等指标。测量时间从早上09:00开始到下午18:00结束,每隔1 h测定1次,种参试植物选择健康、长势均一的树3棵,每棵树选择3片叶,结果取平均值。
2.3 抗氧化酶活性、超氧阴离子自由基及脯氨酸测定

2.4 数据分析与处理
数据分析采用SPSS 23.0完成,其中,方差分析采用多重比较分析中的邓肯检验(Duncan’s test),图表在Microsoft Excel 2010软件中完成。
3 结果与分析
3.1 不同植物的光响应过程特征
4种植物的光响应曲线变化趋势呈现相似性(图1)。在一定的光合有效辐射(PAR)范围之内,净光合速率随着PAR的增加而升高,当PAR增加到一定值后,净光合速率开始缓慢增加并最终趋于平缓。PAR为低于20 μmol·m-2·s-1时,由于光照强度较弱,各植物的Pn基本小于0,此时表明植物的呼吸作用强于光合作用。PAR在100~120 μmol·m-2·s-1时,净光合率上升快慢依次为山黄麻、虾子花、车桑子、天干果。当各个植物的净光合速率达到最大时,对应的PAR也各不同。

图1 不同植物的光合响应曲线
通过光响应参数(表1)可以看出,4种植物的最大净光合速率(Pmax)从大到小依次为:山黄麻、虾子花、车桑子、天干果,说明山黄麻的光合能力最强,天干果的光合能力最弱。虾子花的光饱和点(LSP)为1 236 μmol·m-2·s-1,略高于其他3中植物,说明虾子花拥有较强的强光利用能力,且不易发生光抑制。表观量子效率是植物光能利用效率的体现,研究表明植物的最大初始量子效率理论值为0.08~0.125 μmol·m-2·s-1,但在自然条件下实测值会小于理论值,一般在0.04~0.07 μmol·m-2·s-1[25],本文拟合得出的表观量子效率在0.037~0.085 μmol·m-2·s-1(表1),可以看出山黄麻的叶片光能转化效率高。4种植物的呼吸速率的大小为虾子花、山黄麻、车桑子、天干果。

表1 不同植物光合响应曲线参数
3.2 不同植物环境因子及生理指标日变化
3.2.1 环境因子日变化
从叶片周围环境因子的日变化中可以看出(图2、3),上午09:00—11:00,不同植物间光合有效辐射(PAR)与气温变化相似,呈现出上升趋势,空气相对湿度变化表现为下降趋势;在11:00—16:00,PAR、气温及空气相对湿度表现为上下波动;在16:00后,PAR开始明显下降。PAR在14:00时最大,为1 604.8 μmol·m-2·s-1,气温在15:00时达到最大,为41.8 ℃,空气相对湿度在上午09:00时最大,为41.6%。

图2 不同植物的平均光合有效辐射日变化
3.2.2 净光合速率
如图4所示,4种植物的净光合速率(Pn)均呈现“双峰”型曲线,不同的是山黄麻、虾子花的峰值相较于车桑子、天干果来说更明显。随着时间的推移,Pn显示先增大后降低,然后再增大最后降低的趋势,Pn在11:00、14:00各出现最大值,说明4中植物均有“午休”的现象。4种植物中虾子花1 d中的Pn一直较大,且日变化波动较大,天干果和车桑子一天中的Pn较低,Pn日变化波动也较小。

图3 不同植物的平均气温和相对湿度日变化

图4 不同植物的净光合速率日变化
3.2.3 胞间二氧化碳摩尔分数
胞间CO2摩尔分数(Ci)指植物细胞之间的CO2摩尔分数,能反映出光合作用过程中CO2的耗散与积累,由图5可知,Ci的日变化大体呈现“V”型,即早晚高,中午低。4种植物出现最低Ci的时间点为13:00,在14:00时,4种植物的Ci在间存在略微上升现象,之后,山黄麻和天干果的Ci处于缓慢上升状态,而虾子花和车桑子则先下降然后再上升。
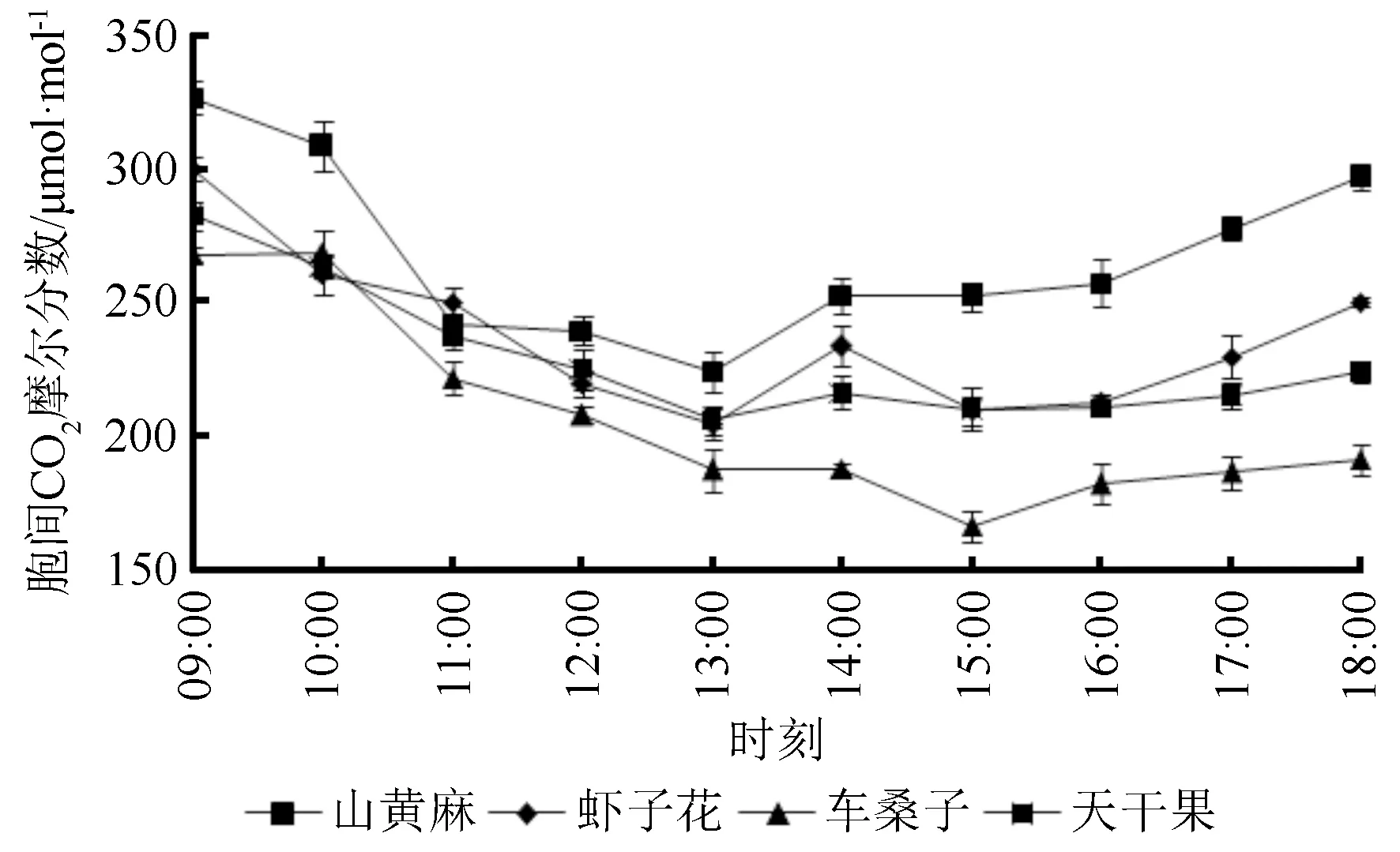
图5 不同植物的胞间二氧化碳摩尔分数日变化
3.2.4 气孔导度
气孔是植物与外界气体交换的主要通道,可控制O2、CO2的交换及蒸腾强度[26],而气孔导度(Gs)则反映了植物在进行光合作用和蒸腾作用时气孔开合的程度。随着时间的推移,气孔导度的日变化呈现先增加后降低再增加最后降低的变化趋势(图6),这和净光合速率的日变化类似,不同的是,除虾子花外,其他3种植物在10:00时,Gs就达到了第一次峰值。另外,在15:00之后,车桑子的Gs略微上升,16:00后又下降,这应该是15:00之后温度降低(图3),植物为保证细胞与大气间CO2平衡,同时减小Ci过低对光合作用的影响,从而调节Gs略微上升。
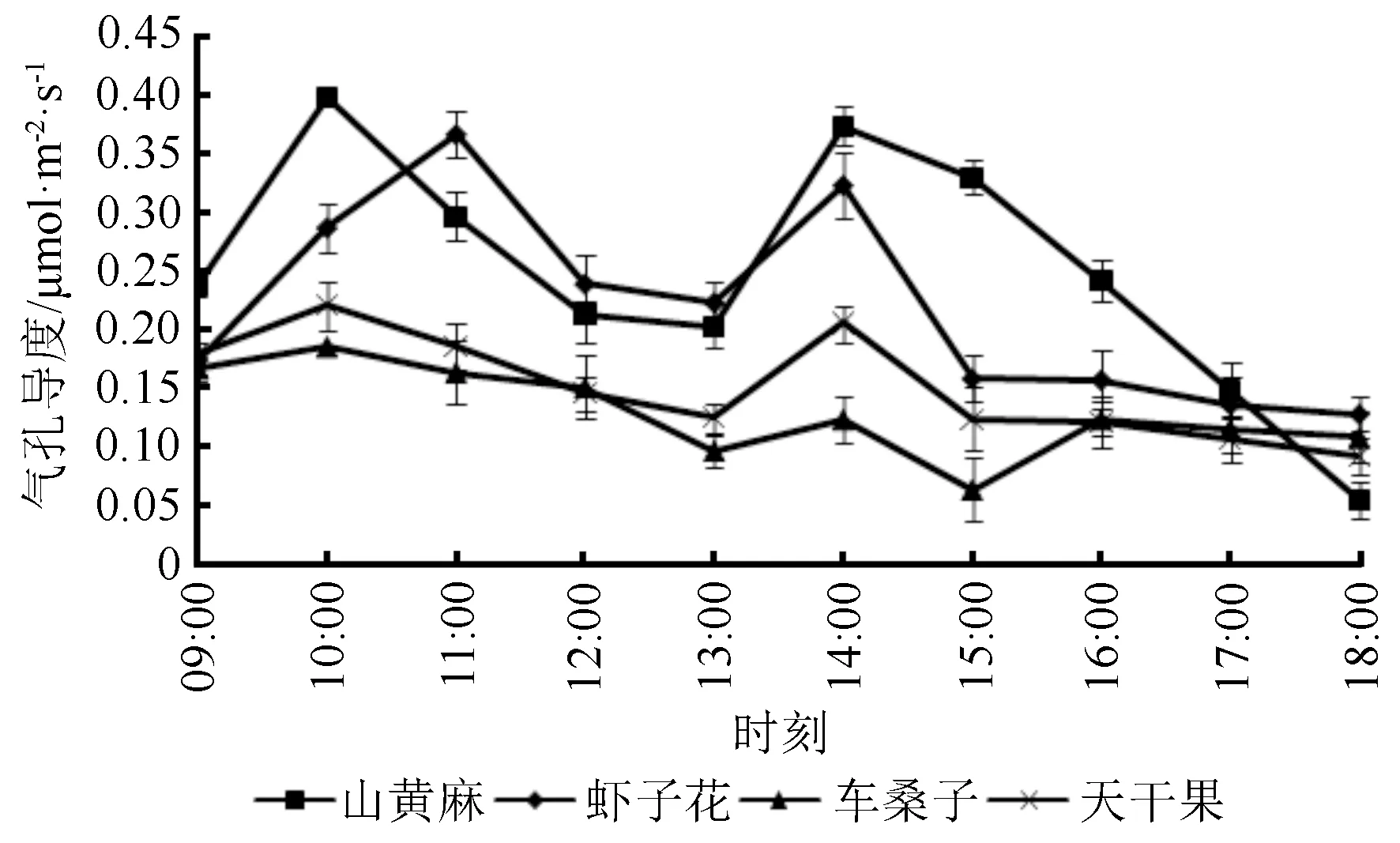
图6 不同植物的气孔导度日变化
3.2.5 蒸腾速率
从图7可以看出4种植物的蒸腾速率(Tr)日变化趋势相同,呈现出先增大后降低再增大最后降低的变化。4种植物Tr均在11:00、14:00达到两次峰值,在达到两次峰值时,山黄麻、虾子花的Tr均大于天干果和车桑子。在中午13:00时4种植物Tr都存在减弱趋势,这是由于干热河谷区中午的PAR和气温较高,植物为避免高蒸腾导致体内散失较多水分从而对胁迫表现出的应对策略。
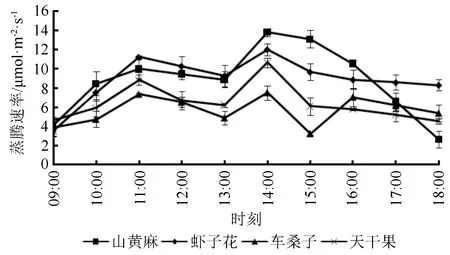
图7 不同植物的蒸腾速率日变化
3.3 不同植物间的叶绿素质量分数
叶片中叶绿素质量分数可决定其对光能的吸收与转换,但随着逆境伤害增加,其质量分数逐步降低,从而使光合速率下降。表2可知,不同树种叶绿素b质量分数之间无显著差异(P>0.05);山黄麻,车桑子与其他植物的叶绿素a质量分数间存在显著差异(P<0.05),其质量分数由大到小依次为山黄麻、车桑子、虾子花、天干果;不同树种间类胡萝卜素质量分数也存在显著差异(P<0.05),其中天干果的类胡萝卜素含量显著低于其他植物(P<0.05)。不同植物的总叶绿素与叶绿素a质量分数变化相同,且山黄麻,车桑子与其他植物的总叶绿素质量分数之间也存在显著差异(P<0.05)。

表2 干旱对不同植物光合叶绿素质量分数的影响 mg·g-1
3.4 不同植物抗氧化酶、脯氨酸及超氧阴离子自由基含量
当植物生长在适宜的环境中,体内活性氧自由基的产生与清除处于平衡状态,在遭遇环境胁迫后,体内产生的自由基将超过植物本身清除的能力,就会引起自由基的积累,并且产生伤害。在红河干热河谷4种植物叶片中SOD、POD、CAT及脯氨酸含量均无显著差异(表3),表明这几种植物在适应干旱条件时,体内均产生较多的SOD、POD、CAT来清除因胁迫产生的活性氧自由基,从而避免植物遭受伤害。通过比较4种植物叶片内的超氧阴离子自由基含量,我们发现山黄麻的超氧阴离子自由基含量最少,且与车桑子、虾子花超氧阴离子自由基含量间存在显著差异(P<0.05)。这说明山黄麻较其他3种植物能更有效清除体内产生的自由基,降低膜脂过氧化作用。

表3 干旱对不同植物抗氧化酶、脯氨酸及超氧阴离子自由基的影响
4 讨论
在植物光合日变化中,净光合速率作为外界环境和植物光合作用的综合反应具有重要的研究价值,环境中的光合有效辐射,温度,相对湿度及植物叶片自身叶绿素质量分数等因素均会影响植物的净光合速率,且这些影响因素是综合表现的[27-28]。此次研究中,4种植物的净光合速率日变化呈现“双峰”,在11:00前后出现峰值,这主要是因为在09:00—11:00时,PAR是限制净光合速率的主要环境因子,当光合有效辐射及气温开始上升,在11:00达到较高时(图6、图7),净光合速率也就出现了第一次峰值;在11:00—13:00,光合有效辐射和气温维持在1d中的较高水平,较高的温度和PAR会对叶绿体的结构造成破坏,加快暗呼吸和光呼吸作用,降低光合作用,从而导致气孔导度降低,减少对CO2的吸收,所以在13:00时不同植物的净光合速率均降低,出现光合“午休”。为避免植物供能不足,在13:00—14:00时植物的部分气孔重新打开以缓解植物体内CO2不足,因此4种植物的Ci和蒸腾速率存在上升现象,净光合速率也出现第2个峰值。
Farquhar et al.[18]认为,净光合速率、气孔导度及Ci均降低,净光合速率的下降为气孔限制,所以可认为4种植物的净光合速率在13:00前后降低是由气孔因素导致。在14:00—15:00时,4种植物经过长时间的蒸腾作用导致植物失水过多,部分气孔开始关闭,从而使气孔导度、蒸腾速率、胞间CO2摩尔分数及净光合速率均出现下降,体现出气孔对植物光合和蒸腾作用的直接影响[26,29]。
植物叶片光合作用产生的有机物是其生长的物质基础,而净光合速率(Pn)可作为植物生产能力的指标,光饱和点(LSP)和光补偿点(LCP)可表现出植物对弱光和强光的利用能力[30-31]。在比较4种植物光响应过程后,可以看出在100 μmol·m-2·s-1之后,4种植物的净光合速率出现较快的增长,这与前人对干热河谷区生长的余甘子、扭黄茅的研究结果类似[32-33],说明在干热河谷区生长的植物在PAR升高的最初阶段,净光合速率均会出现快速的响应。
通过模型对光合参数进行拟合后可知,4种植物的光饱和点为1 082.4~1 236 μmol·m-2·s-1,阳生木本的光饱和点为600~1 500 μmol·m-2·s-1[34],说明4种植物具有阳性植物特征,对强光有较高的利用能力。不同的是山黄麻在1 082.4 μmol·m-2·s-1时就达到了饱和光强,此时其Pmax在4种植物中最大,为16.5 μmol·m-2·s-1,且与其他植物之间存在显著差异(P<0.05),由此推断山黄麻可积累较多的光合产物,其生长代谢可能也高于其他3种植物。天干果拥有较低的光补偿点和较高的光饱和点,说明在红河干热河谷区这一生境下,天干果对光环境有较强的适应性。
非直角双曲线拟合的光响应参数具有生物学意义,且拟合曲线比较符合植物的光响应过程[35]。本文采用非直角双曲线模型拟合光合参数的结果比较理想,能够基本反应各植物的光合特点,4种植物各自在达到饱和光强之后,净光合速率并没有下降,因此,也存在估算的Pmax大于实测值的结果[31,36-37]。
5 结论
虽然红河干热河谷区生长的4种植物对光适应有所不同,但是4种植物叶片中抗氧化酶活性之间无显著差异(P>0.05),说明它们在应对逆境胁迫时均能有效保持植物体内活性氧产生与清除间的平衡,增加抗逆性,避免生物膜遭受膜脂过氧化,表现出对红河干热河谷环境较强的适应性。
