变异链球菌sepM基因中存在与变链素IV形成相关的突变类型
2021-07-08刘姗姗刘玉东张容秀路晓淼
刘姗姗,刘玉东,张容秀,路晓淼,胡 浩,胡 洁,张 凯,孙 钰
蚌埠医学院1第一附属医院口腔科,安徽 蚌埠 233004;蚌埠医学院2组织胚胎学教研室,3卫生检验与检疫学教研室,安徽 蚌埠 233030
龋病是牙体硬组织发生慢性进行性破坏的一种常见的口腔疾病。党和国家一直高度重视人民健康,“健康中国2030”规划纲要中提出“加强口腔卫生,12岁儿童患龋率控制在25%以内”。根据第四次全国口腔流行病学调查结果,我国3~5岁儿童的龋病患病率分别是50.8%、63.6%及71.9%,12~15 岁青少年龋病患病率分别是38.5%、41.2%、43.3%、44.4%;而老年人龋病的患病率更是高达98.1%[1-3]。由此可见,龋病的防治十分重要。
口腔定植菌间的相互作用在龋病的发生中发挥重要角色[4-6]。在众多的口腔定植菌中变异链球菌(Streptococcus mutans)和格氏链球菌(Streptococcus gordonii)是两种研究较多的细菌[7-8]。变异链球菌是重要的致龋菌,其通过代谢口腔环境中的碳源产生酸性产物,导致牙齿脱矿,这种过程持续存在即产生龋病[9-10],而格氏链球菌是定植于牙面的非致龋菌,其代谢产生的过氧化氢可有效抑制变异链球菌生长,产生的碱性产物氨可平衡变异链球菌产生的酸性环境,在龋病的发生中起重要的抑制作用[11-13]。同时,临床研究数据表明,牙菌斑样本中变异链球菌的检出率与龋病呈正相关,而格氏链球菌的检出率与龋病呈负相关[14-15]。因此,变异链球菌与格氏链球菌的相互竞争作用结果是口腔定植菌影响龋病发生的重要因素。
变链素Ⅳ是变异链球菌产生的拮抗格氏链球菌生长、推动龋病发生的一类重要细菌素[16]。研究表明,变链素IV的表达受变异链球菌SepM蛋白的调控,以变异链球菌UA159标准株为参照,sepM敲除株和nlmAB(变链素IV编码基因)敲除株的表型一致,均无变链素Ⅳ的产生,而sepM回补株的变链素IV表达水平与标准株(即野生株)一致[17]。SepM是由346个氨基酸残基编码产生的变异链球菌细胞壁锚定蛋白,主要包括跨膜区(第10~26个氨基酸残基)、PDZ区域(第131~195个氨基酸残基)及C末端(第233~314个氨基酸残基)[17]。由21个氨基酸残基构成的感受刺激肽(CSP-21)可识别ComDE双组份调控系统(ComDE系统),继而激活该系统正调控的变链素IV的表达[18]。基因突变可通过影响基因或蛋白的表达水平进而影响相应的表型,然而变异链球菌SepM的编码基因sepM序列中是否存在影响变链素IV表达的突变类型尚不清楚。因此,本文应用前期牙菌斑样本中分离并保存的变异链球菌临床株,以细菌基因组突变对细菌毒力的影响为出发点,初步探索细菌互作在龋病发生中的机制,为寻找龋病预测和防治的新靶标提供实验依据。
1 材料和方法
1.1 变异链球菌临床株变链素IV形成能力的检测
80株变异链球菌临床株变链素IV的检测参考我们前期的研究,即每株临床株在脑心浸液肉汤培养基(BHI)液体培养基于37℃、5%CO2环境下过夜培养,调A600至0.3。取10 μL菌液于BHI琼脂板上,12 h后将等量的指示菌格氏链球菌(戈登链球菌)ATCC10558 接种于变异链球菌临床株旁并继续培养,12 h后变异链球菌旁抑菌圈的大小即为该菌株的变链素IV形成能力[19]。实验重复3次。
1.2 变异链球菌临床株最小生成树和进化树的构建
实验室保存的80 株变异链球菌临床株已采用Illumina NovaSeq6000 PE150平台进行了全基因组测序,继而采用SOAPdenovo软件进行测序结果的组装,测序结果已上传NCBI[20]。根据核心基因组多位点序列分型美国国家生物技术信息中心(cgMLST)方法采用Perl5.18.2构建这80株变异链球菌临床株的最小生成树;采用TreeBeST软件根据最大似然法构建这80个样本的进化树,步进参数设置为1000;采用Interactive Tree Of Life(iTOL)v5(https://itol.embl.de/)绘制进化树[21-23]。
1.3 变异链球菌临床株sepM基因突变的分析
采用GeneMarkS 软件(verison4.17)(http://topaz.gatech.edu/)对变异链球菌临床株的最终组装结果进行编码基因预测及过滤[24]。以变异链球菌UA159菌株的全基因组序列(NC_004350.2)为参考,将预测得到的基因信息与该参考基因组进行比对,将CDS 区cover≥70%并且identity≥50%的预测基因定义为已知基因[25]。将变异链球菌临床株sepM基因与参考基因组sepM全长序列进行比对,分析临床株中sepM的突变类型。
1.4 统计学分析
采用IBM SPSS20.0软件进行统计分析。定性资料的分析和比较采用pearson chi-square检验(理论频数≥5)、连续校正法[1≤理论频数<5(单元格数≤20%)]或Fisher精确概率法(不符合前两者的情况);定量资料以均数±标准差表示,其分析和比较采用t检验,定量资料中每组样本量不足3的未纳入统计分析。P<0.05认为差异具有统计学意义。
2 结果
2.1 变异链球菌临床株最小生成树和进化树的构建
80株变异链球菌株共有25株细菌有变链素IV形成能力(变链素IV形成组),另55株细菌缺乏变链素IV形成能力(无变链素形成组)。根据变异链球菌临床株的核心基因组数据,我们构建了这两组菌株相应的最小生成树(图1)和进化树(图2)。
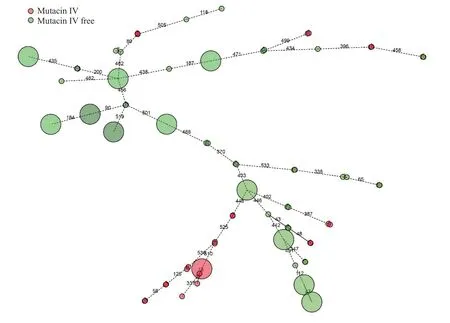
图1 80株变异链球菌临床株的最小生成树Fig.1 Minimum spanning tree of the 80 clinical isolates of S.mutans.
2.2 变异链球菌临床株sepM基因突变的分析
80株变异链球菌临床株的基因组与参考基因组UA159比对后,均存在已知基因sepM。临床株中sepM基因均全长表达,不存在碱基的插入和缺失,并且共检测到34个点突变。在这些点突变里,包含26个同义突变和8个错义突变(表1)。
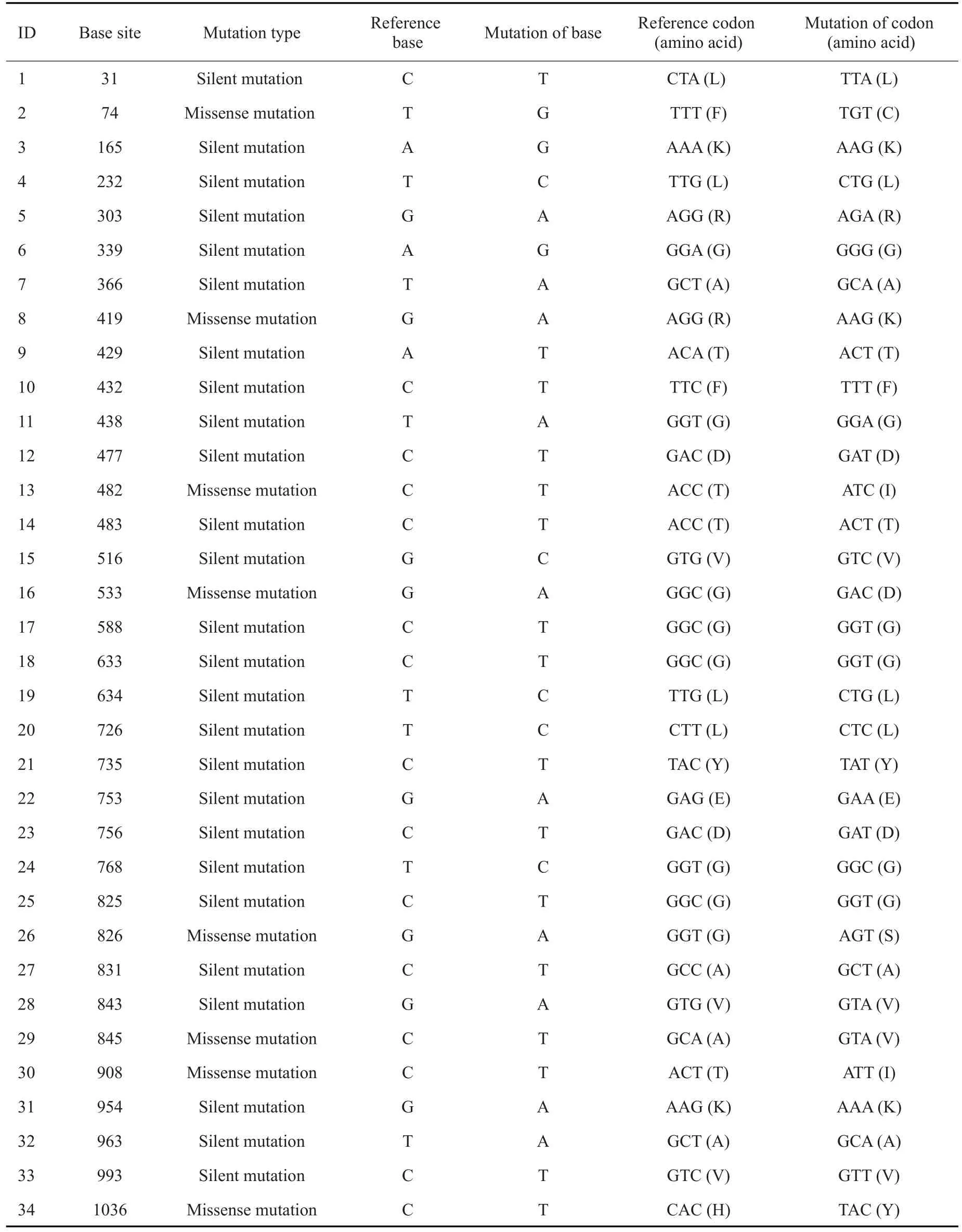
表1 变异链球菌临床株中sepM基因突变类型Tab.1 Mutations of sepM in 80 clinical isolates of S.mutans
2.3 sepM基因突变与变链素IV形成能力关系的分析
变链素IV形成组和无变链素IV形成组间差异分布的sepM临床点突变共4个,其中错义突变和同义突变位点各占50%。这4 个突变位点(C31T、G533A、C756T、C1036T)在变链素IV形成组的分布频率显著高于无变链素IV形成组。这些差异分布的突变位点进一步的分析结果表明,突变组变链素IV的形成能力也显著高于非突变组(表2)。
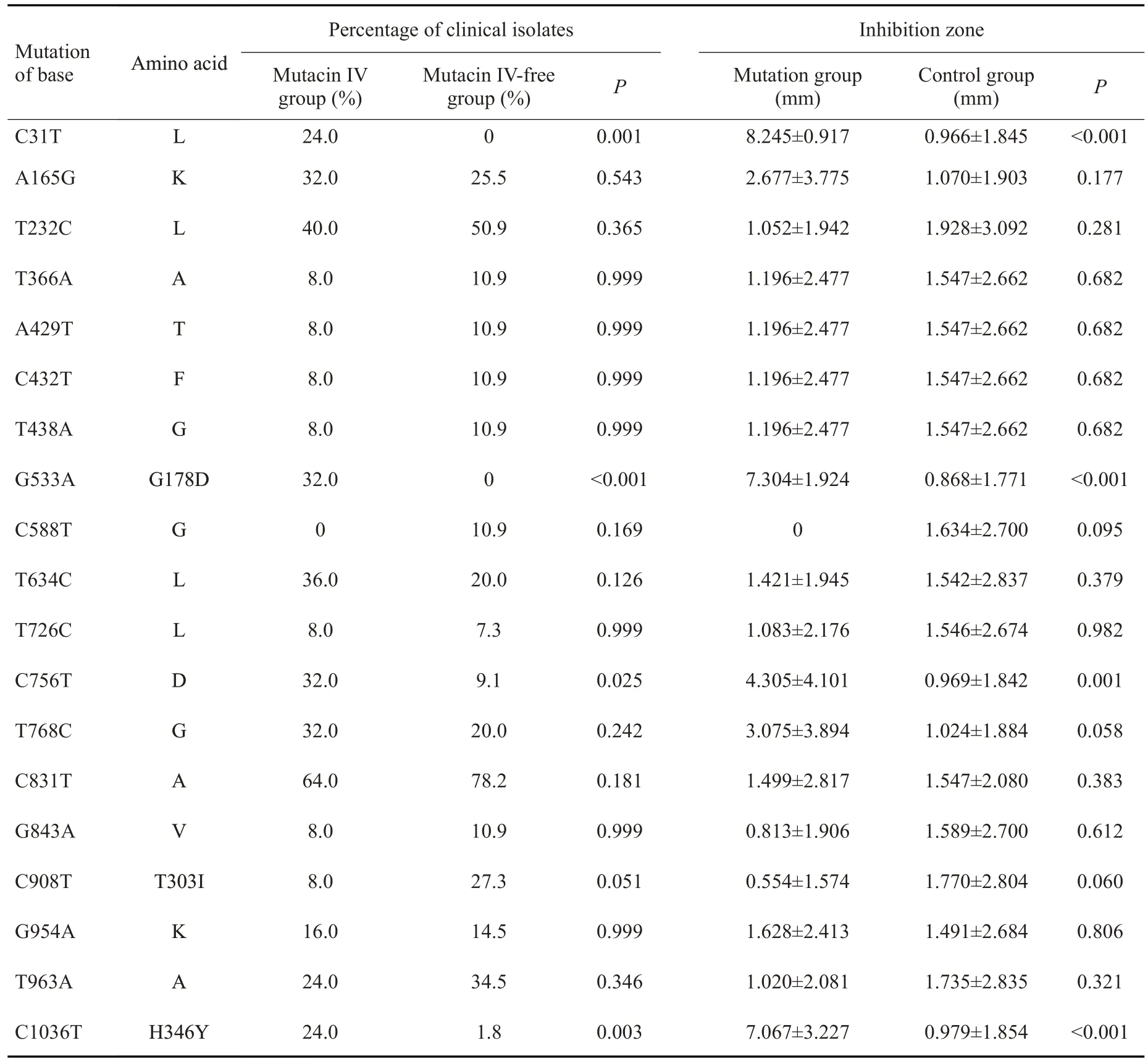
表2 80株变异链球菌临床株中差异分布的sepM基因突变类型与变链素的关系Tab.2 Association of the differentially distributed sepM mutations with mutacin IV production in the 80 clinical isolates of S.mutans
3 讨论
最小生成树主要通过菌株间等位基因的区别来了解菌株间的差异,而进化树是分析细菌起源的重要手段,这两种方法已广泛用于细菌基因组的分析[26-30]。多位点序列分型法(MLST)是既往用于变异链球菌流行病学研究的重要分型方法[31-35]。我们前期研究发现,该方法难以区分基因组相似的细菌,因此,我们构建了优于MLST的cgMLST[20]。基于该方法中的核心基因组,并结合本研究中这些菌株的变链素IV的表达水平,我们构建了这80株细菌的最小生成树和进化树。本研究发现,变链素IV形成组内部菌株之间的差异少于组间菌株之间的差异。这可能与变链素IV形成组组内菌株的起源相似或相同相关。因此,我们进一步构建这80株细菌的进化树,而进化树的结果也与最小生成树保持一致,即在最小生成树中等位基因差异小的菌株在进化树中均起源自相同的分支。
以往研究报道,细菌基因突变可能通过改变自身毒力的表达进而增加或降低机体的患病风险。例如,金黄色葡萄球菌WalK蛋白的223位氨基酸G突变为D后,导致WalK 和下游WalR的磷酸化水平下调,从而降低了金黄色葡萄球菌的细胞壁厚度、自溶能力及其对万古霉素的敏感性[36];侵袭性肺炎链球菌源性脑膜炎患者中分离的肺炎链球菌细胞壁代谢相关蛋白PBP1b的编码基因pbp1b的641位碱基有24%的A突变为C,显著高于对照组(11%);进一步在肺炎链球菌R6标准株中构建pbp1b A641C 突变株,发现突变株的最小杀菌时间显著长于标准株[37]。因此,本文应用与疾病发病状态更接近的变异链球菌临床株,在了解有变链素IV形成能力和无变链素IV形成能力的变异链球菌临床株之间的差异和进化趋势后,进一步探索sepM突变与变链素IV形成的关系。
CSP-21是变异链球菌产生的并分泌至菌体外的由21个氨基酸残基构成的多肽,其末端3个氨基酸可被剪切形成由18个氨基酸残基构成的多肽CSP-18,CSP-18是比CSP-21活性显著的多肽,其进一步与下游ComD结合,促进ComD及其下游ComE的磷酸化,磷酸化的ComE识别并结合至变链素IV编码基因nlmA和nlmB的启动子区,激活变链素IV的表达[18,38-39]。而SepM是裂解CSP-21生成CSP-18的关键蛋白酶[17]。本研究发现,虽然所有菌株均能检测到完整的1041bp sepM序列,但是这些菌株的变链素IV水平却存在区别。这也进一步提示并初步印证变链素IV的表达差异可能与sepM基因存在多态性相关。sepM序列比对结果亦表明,变链素IV形成组和无变链素IV形成组间存在4个差异分布的sepM临床突变位点,并且这些突变位点的突变组和非突变组菌株中变链素IV水平也存在显著差异。这提示,sepM基因突变可能与变链素IV的形成相关。后期需要更多的样本量并结合点突变的功能实验进一步验证具体的有意义的位点在变链素IV形成中的作用。
综上所述,变异链球菌临床株中可能存在与变链素IV形成密切相关的点突变,这为后续针对致龋菌的毒力干预提供了理论依据,并为龋病的精准预防提供了潜在的方向。
