陇南山区黄瑞香天然种群实生苗更新研究
2021-07-05张春云王勤礼王建东王海平张春梅
闫 芳, 张春云, 王勤礼, 王建东, 王海平, 徐 涛, 张春梅,, 陈 叶, 朱 高
(1. 河西学院祖师麻原植物应用技术研究所, 甘肃 张掖 734000; 2. 甘肃泰康制药有限责任公司, 甘肃 武威 733000;3. 甘肃省河西走廊特色资源利用重点实验室, 甘肃 张掖 734000; 4. 肃南明海林场, 甘肃 张掖 734000)
陇南山区位于甘肃省东南部,地处青藏高原东部,呈高山峡谷、盆地河谷交错分布、沟壑纵横的复杂地形地貌,以褶皱带发育的高低山地与盆地而得名。山区地势东低西高,光热充足,最低海拔和最高海拔相差3 500 m左右[1],属亚热带湿润气候向西北高寒湿润气候的过渡地区,在垂直方向产生北亚热、暖湿、温和、湿凉、冷凉5种气候带。因此,此区植被种类丰富,大致可分为落叶阔叶林、针阔混交林、亚高山针叶林及高山灌丛草甸等,其中种子植物4 000多种、药用植物469 种[2]。黄瑞香(DaphnegiraldiiNitsche.)为瑞香科瑞香属的落叶小灌木,胸径40~80 cm,植株高40~100 cm,分布在此区海拔2 000~2 600 m的山坡灌丛、林缘、沟谷地带。黄瑞香的茎皮即西北地区的民间药“祖师麻”,此药在甘肃省民间自产自用历史已久,主治风湿痹痛、四肢麻木、头痛、肝区痛等多种痛症和跌打损伤等症[3-5]。该药材来源目前依靠野生采挖,多年来连根掘起的掠夺性采药方式已致使各道地产区面临野生黄瑞香种质资源枯竭的现状[6],因此,对黄瑞香进行有效地种群抚育和恢复是目前的主要研究任务。
林木种群实生苗更新是林木利用自身繁殖能力形成新一代幼苗的复杂生态过程,主要受到地形、海拔、土壤条件等环境因子的影响以及林分盖度、灌草本植被盖度等生境因子的影响[7]。黄朗等研究表明灌木盖度、草本盖度、坡位和坡向是影响湖南栎类(Quercusspp.)天然次生林幼树更新的主要因子[8];陈本学等认为白藤(CalamustetradactylusHance)较低的幼苗转化率是制约其幼苗天然更新的关键环节[9];任学敏等研究发现土壤全磷含量、坡向、坡度和有机质含量是影响锐齿槲栎(Quercusalienavar.Acuteserrata)更新密度的主要环境因子[10];李宁等指出存在阶段性变化的生境是引起红豆杉(TaxuschinensisPilger Rehd.)幼树数量远少于幼苗这一衰退现象的主因[11]。实生苗更新作为植物群落生态系统自我繁衍和恢复的主要方式,其生态学过程本身持久而复杂,且更新树种和生长环境不同,实生苗天然更新的关键影响因子亦各不相同。因此,了解种群实生苗生长现状,研究各影响因子与植株实生苗更新之间的关系和探究实生苗更新的关键影响因子对林木种群恢复具有重要的理论与实际意义。在黄瑞香人工繁育研究方面,种子休眠机理和资源调查等方面研究已有学者报道[12-14],但野生黄瑞香种群天然更新相关研究较少。本文旨在通过研究陇南山区不同生境黄瑞香实生苗生长发育及其与环境之间的关系,探索该区域黄瑞香野生种群自然更新的影响因素和规律,为陇南山区黄瑞香的种群恢复和持续经营提供科学依据。
1 材料与方法
1.1 研究区概况
本研究区位于甘肃省陇南市武都区马营镇(104°51′16″E,33°34′53″N)及周边地区。该区域地处陇南市武都区西北郊,海拔2 100~2 500 m,年降雨量600~800 mm,土壤以肥沃、性热而紧的黄棕土壤为主。区域内山峦起伏,西部山高谷峡,无成片森林,有零星树木,杂草丰茂,自然覆盖较好,大片黄瑞香野生群落很少见到,大多是以黄瑞香为主的灌木林和疏林,常以斑块状分布在半阳坡和半阴坡半生境。伴生植物由大火草(Anemonetomentosa),阴坡高山柳(SalixCupularis),阴坡直穗小檗(Berberisdasystachya),野李子(prunusselicina),野艾蒿(Artemisialavandulaefolia),甘肃蒿草(Kobresiakasuensis),迷果芹(Sphallerocarpusgracilis),长茎柴胡(Bupleurumiongicaule),麻花艽(Gentianastraiminea),紫野燕麦(Hordeumviolaceum),喜荫悬钩子(Rubusmesogaeus),多节雀草(Bromusplurinodis),双叉细柄毛(Ptilagrostisdichotoma),党参(Codonopsispilosula),长芒草(S.bungeana),披碱草(Elymusdahuricus)等植物组成。
1.2 样地设置
以陇南市武都区马营镇马营村清水沟南山和西山的黄瑞香天然种群为对象,于2019年7月在3个分布区的半阴坡和半阳坡设置3块大小为40 m×80 m的调查样地,采用相邻网格法把每个样地划分成20 m×20 m的固定样地,总数24块,样地基本情况见表1。固定样地的黄瑞香树龄相近,植株发育较好,由于山形陡峭,故未进行规模性人为破坏活动。
1.3 样地调查
1.3.1灌草调查 2019年8月对各样地沿坡植物进行林分特征和郁闭度的调查工作,每块样地各设置5个5 m×5 m的灌木样方和1 m×1 m的草本样方,分别在灌木样方内和草本样方内调查各种灌木和草本类植物的种类、数量、高度和盖度,同时调查样方内枯枝落叶层的厚度。
1.3.2环境因子调查 用罗盘仪(DQY-1A)测定样地的坡向、坡度;用手持GPS定位仪(G128BD)测定样地地理坐标和海拔;空气温度和湿度测定分别用温湿数据采集器(U23-002),光照强度测定用光照计(ZDS-10)。测定时间选择晴朗的天气,在不同生境的固定位置,测定距地表以上0.5 m处的空气温度、湿度及光照强度,测定时间为10∶00~16∶00,测定时间间隔为2 h。样地土壤含水量采用烘干(105℃)法测定,于5个连续的晴朗天,取各样地沿对角线0~20 cm处的土壤用于测定样地土壤含水量。采用国家林业行业标准《森林土壤分析方法》测定土壤养分含量,包括土壤有机质含量,土壤速效氮、磷和钾的含量[15]。
1.3.3实生苗调查 2019年8月中旬至下旬,在各分布区每个样地中的3个灌木样方中,仔细寻找<10.0 cm的黄瑞香实生苗,统计样方中实生苗数量,并测量基径、冠幅、高度和一级分枝数量。基径是通过测定黄瑞香根颈部地面处主茎的直径而得;冠幅为黄瑞香实生苗南北和东西方向宽度的平均值。合并同一生境的调查数据,算出样区内黄瑞香实生苗的平均密度、平均基径、平均高度、平均冠幅和平均分枝数。黄瑞香实生苗个体树龄可通过主茎上的芽鳞痕和茎干颜色(新生枝条呈红棕色)确定,采用SPSS 19.0 软件将各生境中测量的实生苗基径和树龄的原始数据进行拟合,得到黄瑞香不同生境中实生苗基径与树龄的关系:
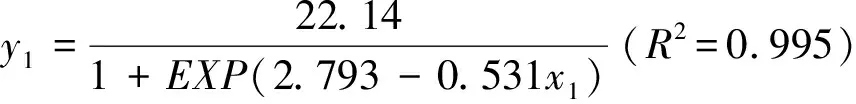
式中:x1和x2分别为各样地半阴坡和半阳坡黄瑞香树龄,y1和y2分别为各样地半阴坡和半阳坡黄瑞香基茎,EXP为欧拉数,样本数均为n=36。套用以上公式计算不同树龄黄瑞香实生苗基茎对应数值,经过多样本的实际测量值和卡方检验(P<0.05),此方程符合实际要求。
1.4 干物质测定
在各样地分别选取接近1至5年生苗龄平均高度、冠幅和基径的黄瑞香实生苗作为标准株,每个苗龄选4株,整株挖下,去掉表面泥土,带回实验室清洗干净,分为根、茎、叶3部分,将叶片用CanoScan LiDE 700F 扫描仪扫描成图片,用Image J软件统计并记录叶面积[16]。将所有以上植株各部位在80℃条件下烘干后测定含水率,据此计算各器官的干质量和总干重[17],最后合并相同条件下的实生苗干质量数据并计算平均值。计算公式如下[18]:
RSR(根冠比)=RB(根干重)/AB(地上部分干重)
LBR(叶干质量比率)=LB(叶片干质量)/TB(总干重)
SBR(茎干质量比率)=SB(茎干重)/TB(总干重)
RBR(根干质量比率)=RB(根干重)/TB(总干重)
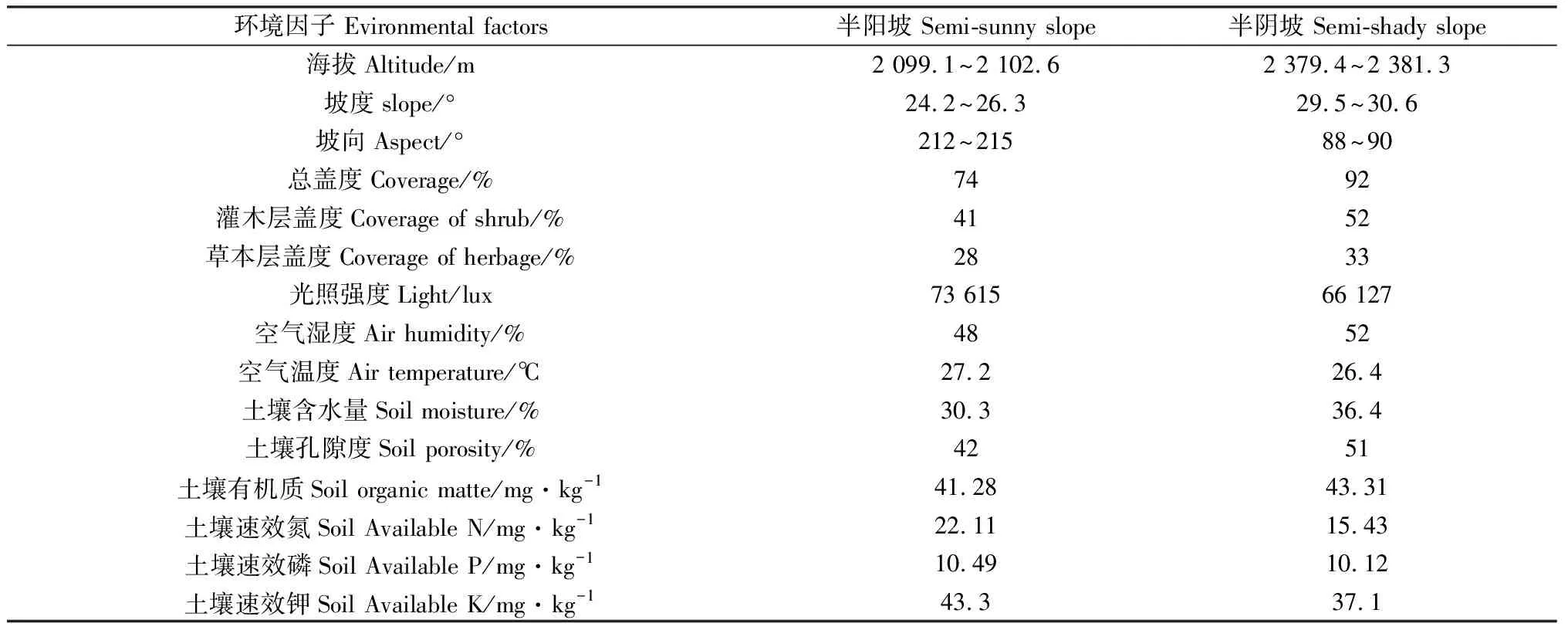
表1 黄瑞香实生苗监测样地基本情况Table 1 Sample plots of D. giraldii populations in different habitat
1.5 数据分析
所有数据用SPSS 19.0软件进行处理,使用One way ANOVA进行单因素方差分析,用Duncan法进行多重比较,最小显著差法(LSD)在0.05 水平确定各个平均值之间的差异显著性。用Origin8.5绘图,用相关性分析方法分析幼苗定居与环境因子的关系。
2 结果与分析
2.1 不同生境黄瑞香实生苗幼苗转化和年龄组成
半阳坡、半阴坡均有1年生黄瑞香实生苗,从单位面积样地内的实生苗数量看(表2),半阳坡生境的黄瑞香实生苗数量为9.17 株·(100 m2)-1,显著高于半阴坡生境(P<0.05)。半阳坡、半阴坡生境下黄瑞香种子幼苗转化率均较低,且差异显著(P<0.05),表现为半阳坡(31.2%)>半阴坡(26.6%)。

表2 不同生境黄瑞香种子幼苗转化率Table 2 Ratio of D. giraldii seedlings to fruit in different habitat
如图1所示,黄瑞香实生苗库总体较小,且各生境黄瑞香实生苗的数量随树龄增长而显著减少(P<0.05),1至5年生黄瑞香实生苗数量表现为半阳坡显著多于半阴坡(P<0.05),半阳坡和半阴坡1年生黄瑞香实生苗的数量占总体数量的28.31%,而5年生的实生苗则只占总体数量的7.55%。其中,半阳坡1至5年生黄瑞香实生苗数量分别占总体数量的29.03%,25.81%,22.58%,16.13%,6.45%,而半阴坡1至5年生黄瑞香实生苗数量分别占总体数量的27.27%,22.73%,22.73%,18.18%,9.09%。
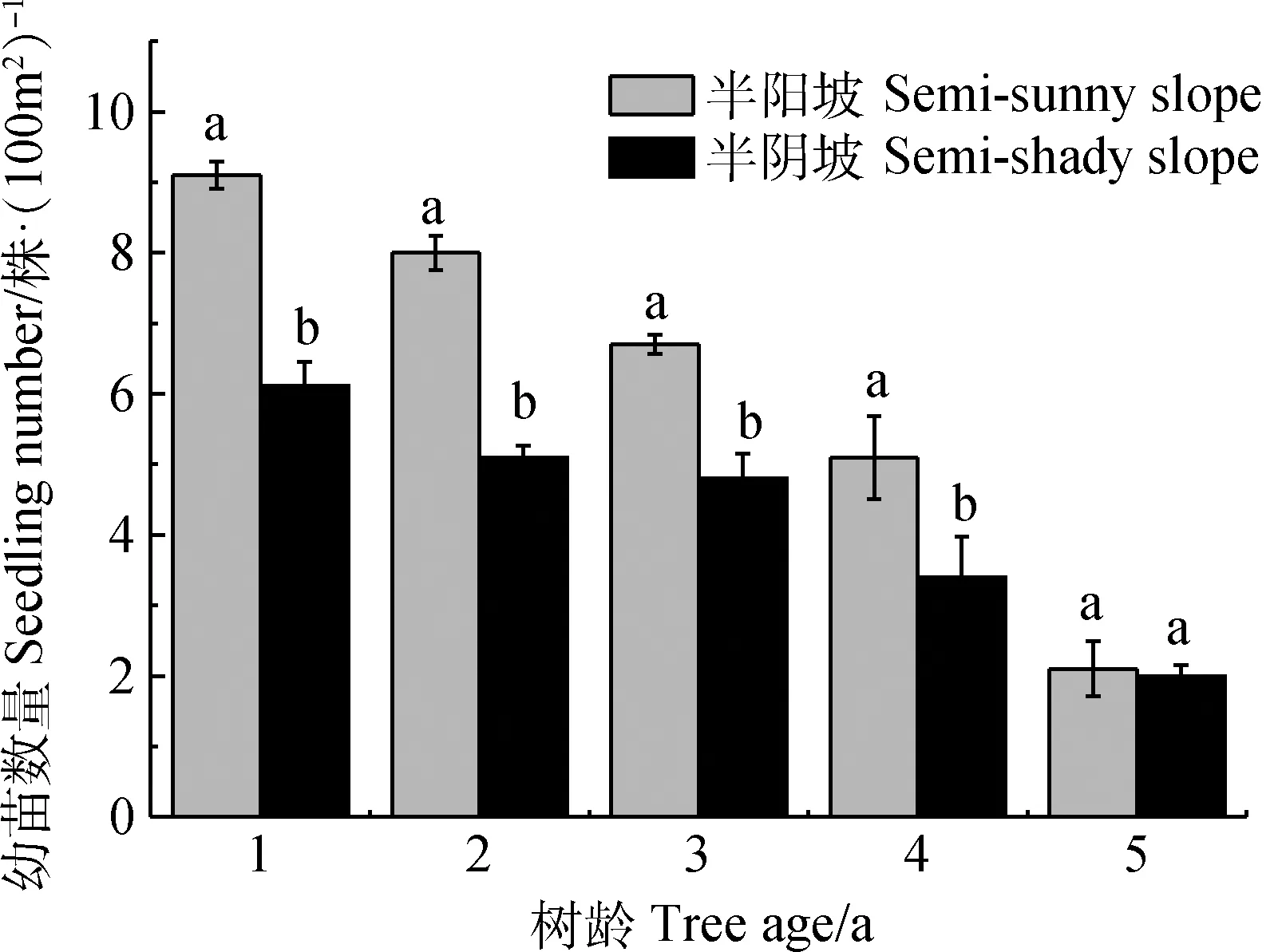
图1 不同生境下黄瑞香实生苗的年龄组成Fig.1 Age structure of D. giraldii seedling in different habitat注:不同小写字母表示分布区间差异显著(P<0.05)。下同Note:Different lowercase letters meant significant difference among different distribution regions at the 0.05 level.The same as below
2.2 黄瑞香实生苗的生长发育特征
陇南山地黄瑞香生育期为160 d,4月上旬返青,9月底停止生长,种子落入草丛深处休眠。由表3可知,黄瑞香在6月初进入初花期,此时生长速度最慢,为0.14 cm·d-1,每个植株平均生长花序16~28个。盛花期生长速度逐渐上升,为0.88 cm·d-1,到7月中旬的结实期,日平均生长速度逐渐下降,为0.23 cm·d-1,7月中下旬进入果实转色期和成熟期时,生长速度基本没有变化。总体看来,黄瑞香实生苗生长速度表现为半阳坡大于半阴坡。
2.3 实生苗的形态特征
随着黄瑞香树龄的增长,各生境实生苗的冠幅、株高、叶面积、主根长、分枝数、茎粗、根幅均逐年增加。其中叶面积、主根长和茎粗呈稳定增加的趋势,主根长和叶面积表现为半阴坡>半阳坡,而冠幅、株高、根幅、茎粗则总体表现为半阳坡>半阴坡。增长趋势具体表现为1年生和2年生实生苗增长缓慢,且两个生境差异不显著,3年生起增长较快,随着树龄的增长,不同生境黄瑞香实生苗生长差异愈加显著。黄瑞香1年生和2年生实生苗不分枝,3年生起开始分枝,株高在此时也开始有较大增幅。主根长/株高从3年生开始明显降低,而根幅/冠幅则呈先缓慢降低后缓慢增加的趋势。从上述黄瑞香实生苗的生长特征可以看出,半阳坡生境较半阴坡生境更有利于实生苗的生长发育。
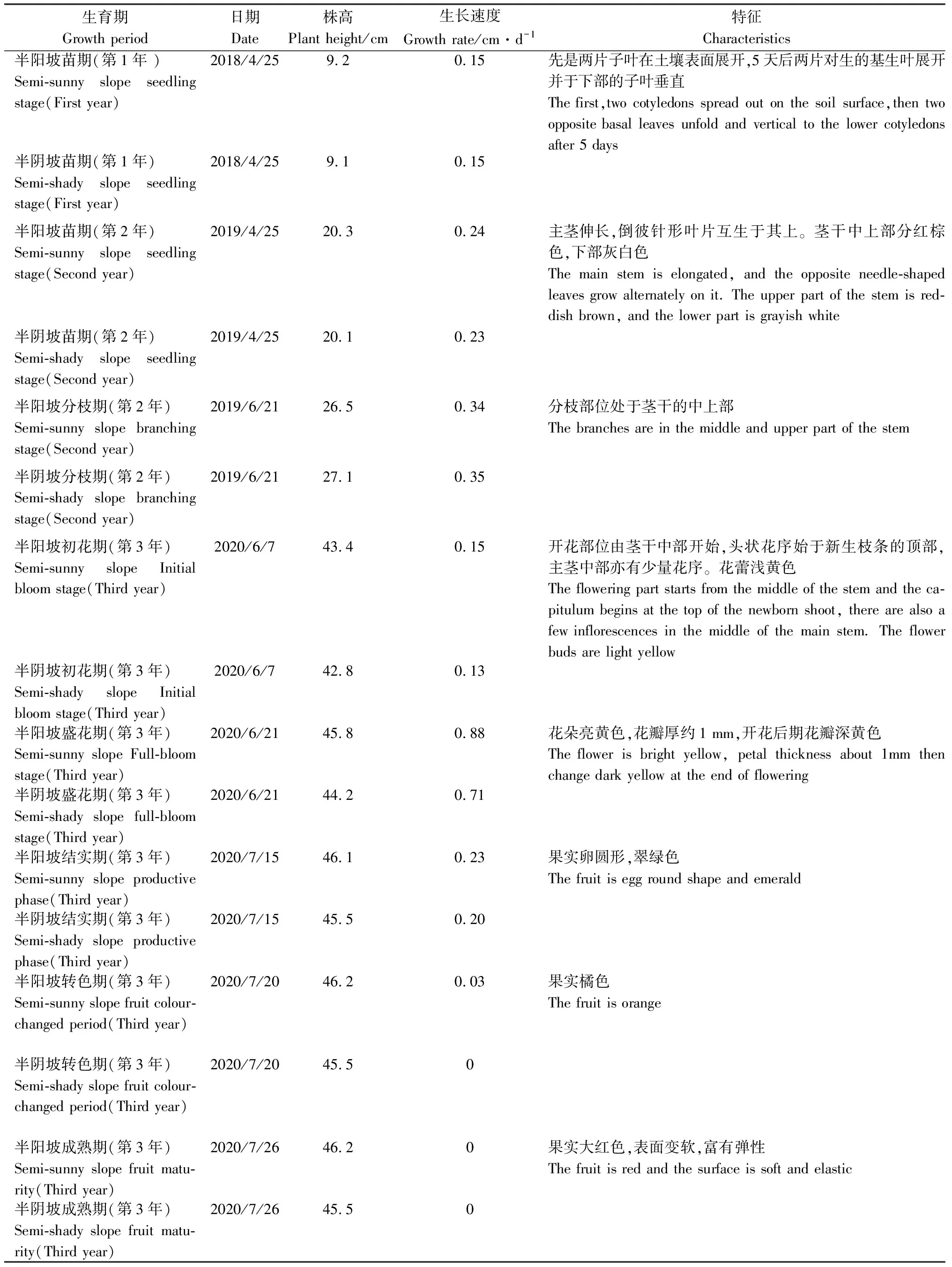
表3 黄瑞香实生苗生长发育观测表Table 3 Observation on growth and development period of D. giraldii seedling
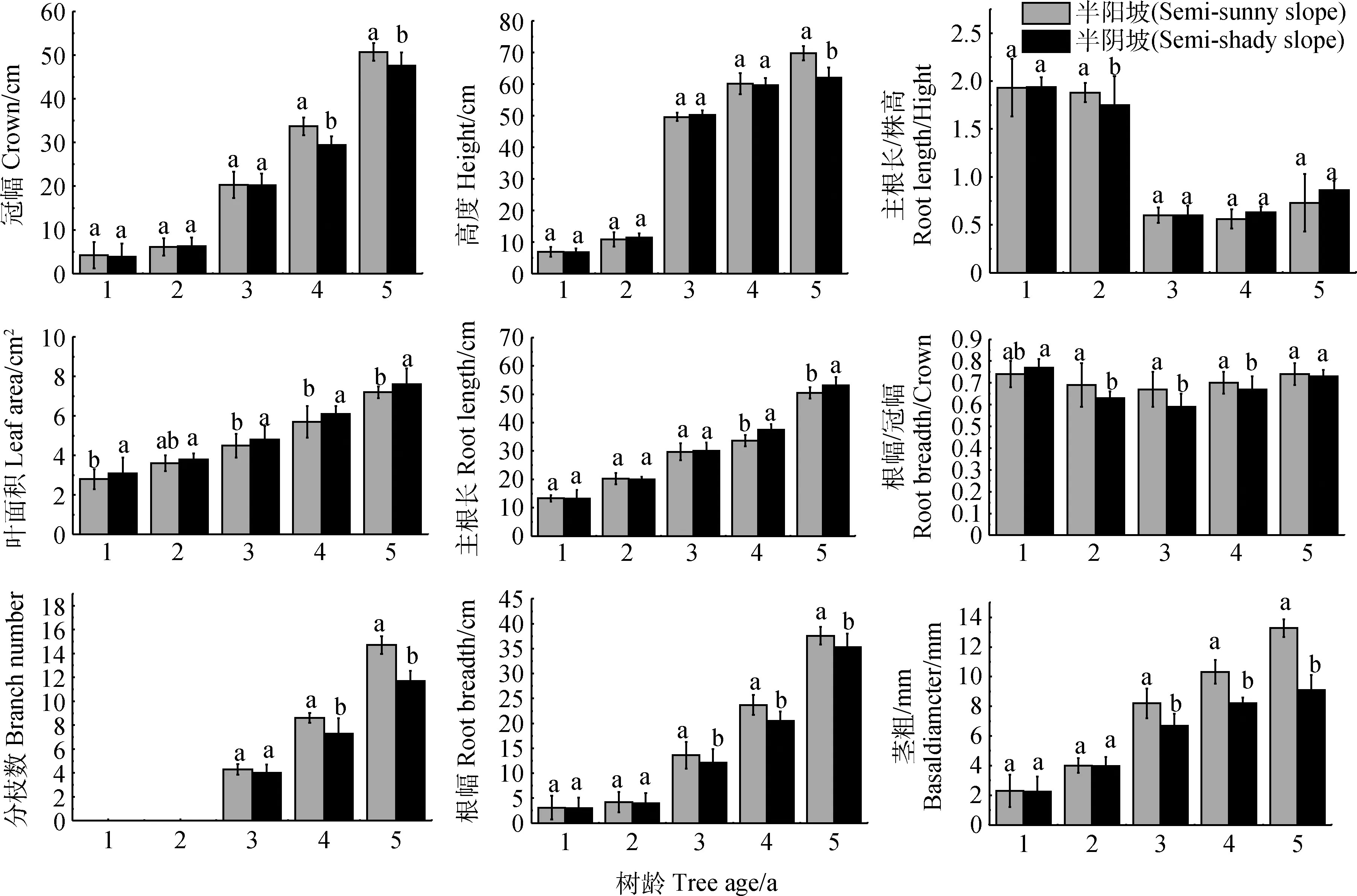
图2 不同生境条件下黄瑞香实生苗的形态特征Fig.2 Morphological characteristics of D. giraldii seedling in different habitat
2.4 黄瑞香实生苗干物质积累及分配
植物干质量的积累反映了植物对环境条件的适应状况[19]。由表4可知,随着黄瑞香树龄的增长,黄瑞香实生苗地上、地下干质量呈显著增加的趋势(P<0.05),除半阴坡1年生幼苗地上干质量略大于半阳坡外,其余黄瑞香实生苗地上、地下干质量均表现为半阳坡>半阴坡。不同树龄黄瑞香实生苗地上干质量均大于地下干质量,说明该植物将更多的资源投入到了地上部分的生长。不同生境条件下黄瑞香实生苗根冠比有所不同,半阳坡和半阴坡实生苗根冠比均呈先降低后增加的趋势,半阳坡生境条件下1年生幼苗根冠比最大,而半阴坡则表现为5年的实生苗根冠比最大,不同生境各树龄的根冠比均表现为半阳坡>半阴坡(表4)。
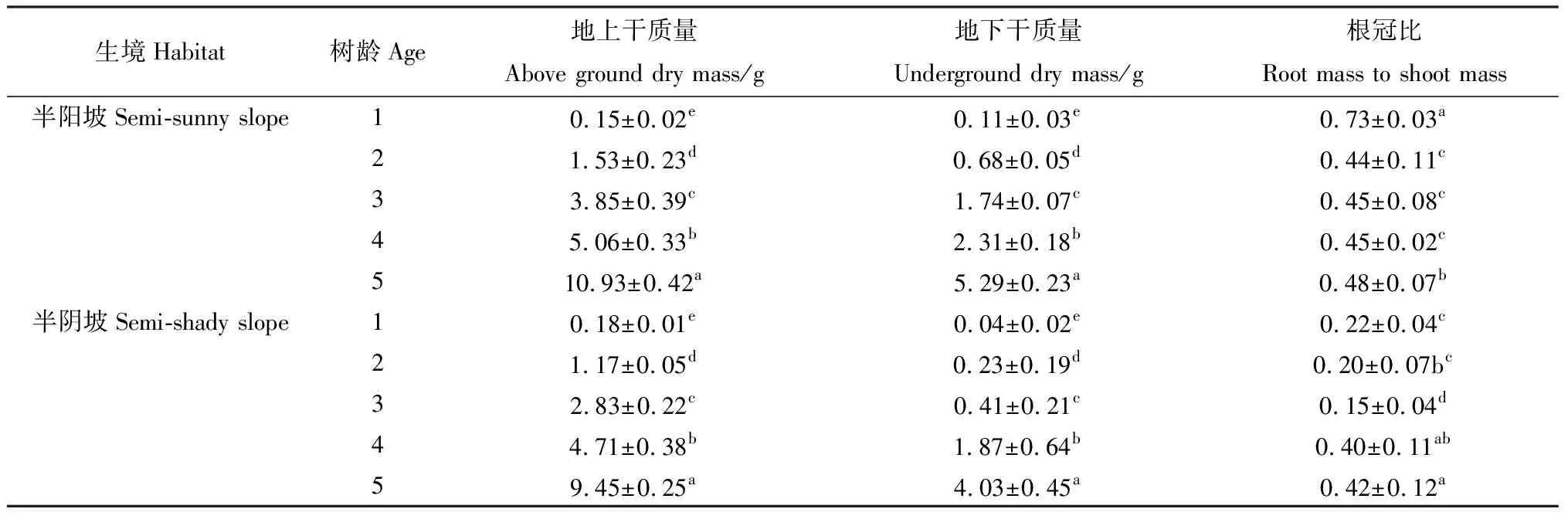
表4 不同生境条件下黄瑞香实生苗干质量分配Table 4 Biomass allocation of D. giraldii seedling in different habitat
不同生境黄瑞香1年生实生苗的茎干重和根干重差异均不显著,不同生境1~3年生实生苗的叶干重差异也不显著。2~5年生实生苗茎干重和根干重、4~5年生叶干重均表现为半阳坡显著大于半阴坡生境(P<0.05),且差异随着树龄的增加而逐步扩大(图3)。5年生半阳坡黄瑞香实生苗茎干重比半阴坡增加了35.8%,叶干质量增加了11.4%,根干质量增加了31.3%。由图3可知,不同生境黄瑞香实生苗各器官的干质量比率表现差异较大。随苗龄的增长,茎的干质量比率呈逐渐增高的趋势,且差异越来越显著(P<0.05);叶的干质量比率呈逐渐下降的趋势,且随着树龄的增大差异越来越小;根的干质量比率则表现为半阳坡各树龄实生苗均不显著,半阴坡的1~3年生实生苗显著降低,随着树龄的增大差异越来越不显著。总体来说,茎和根的干质量比率大致表现为半阳坡>半阴坡,而叶干质量比率则表现为半阴坡>半阳坡。
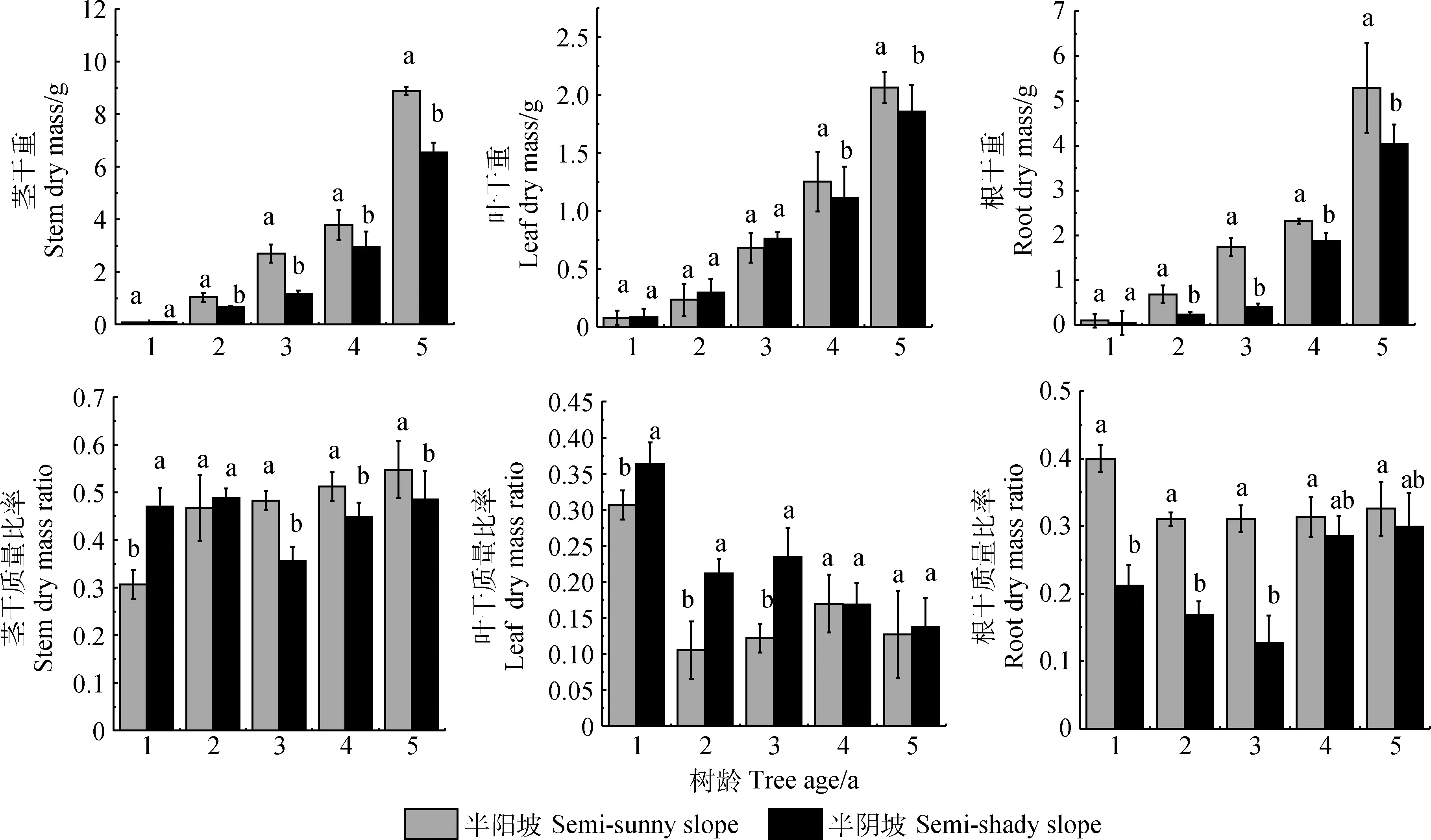
图3 不同生境黄瑞香实生苗各器官干质量Fig.3 Modular dry mass and its ratio of D. giraldii seedling in different habitat
2.5 环境因子对黄瑞香实生苗生长的影响
将野生黄瑞香种群各样地的海拔、土壤有机质、土壤速效氮、土壤速效磷、土壤速效钾等环境因子和总盖度、灌木层盖度、草本层盖度等生境因子作为自变量,将黄瑞香实生苗冠幅、高度和根茎干重作为因变量进行相关分析(n=10),结果如表5所示。实生苗冠幅、高度和根茎干重与总盖度、灌木层盖度、草本层盖度、海拔呈负相关,其中,总盖度对实生苗高度和冠幅的影响分别达到极显著(P<0.01)和显著水平(P<0.05),灌木层盖度对实生苗冠幅的影响达到显著水平(P<0.05)。实生苗冠幅、高度和根茎干重与土壤有机质、速效氮、速效磷和速效钾含量呈正相关,其中,土壤速效氮分别对实生苗高度和冠幅的影响达到极显著水平(P<0.01)和显著水平(P<0.05)。
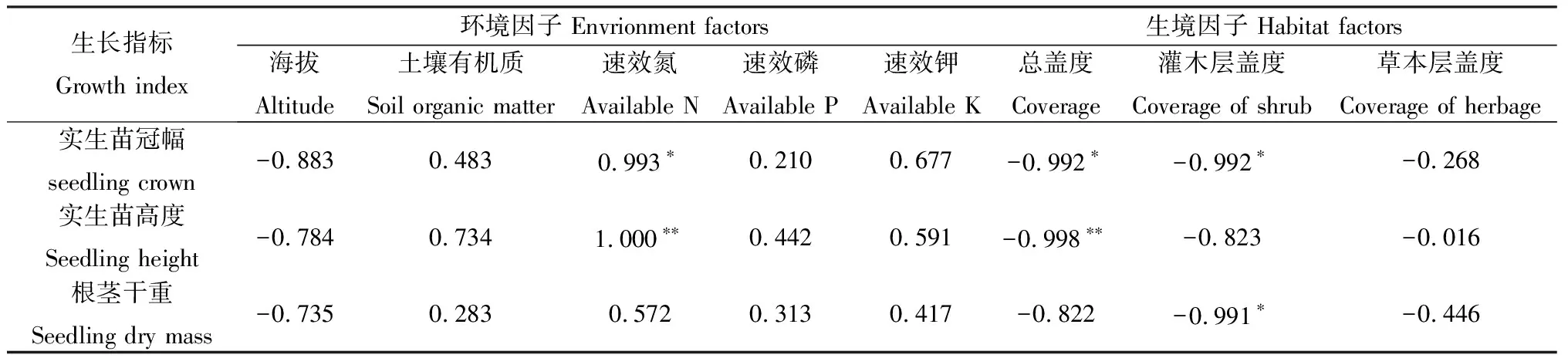
表5 黄瑞香实生苗生长与影响因子的相关性分析Table 5 Correlation analysis bewteen D. giraldii seedling growth and impact factor
3 讨论
植物野生种群的实生苗更新对于维护该物种的群落稳定性来说是一个关键环节,直接影响着该种群的群落组成与结构[20],而种子转化成幼苗这个高风险的过程则成为种群生活史中最为脆弱的阶段[21]。本研究结果显示,从半阳坡生境到半阴坡生境,黄瑞香实生苗密度、幼苗转化率和生长速度均变低,说明半阴坡生境的光照、土壤水分和养分、空气温度等因子可能影响了实生苗的生长发育和定居。两种生境黄瑞香1年生实生苗数量只占幼苗总体数量的28.31%,说明种子在野外自然解除休眠后萌发直至发育成苗这一过程是黄瑞香种群天然更新的一个主要限制因素,而随着树龄的增长,黄瑞香实生苗数量逐渐减少,说明环境因素的干扰影响了黄瑞香小龄苗向大龄苗的转化过程。植物种群的更新过程在不同阶段会受到不同因子的影响,使得实生苗受到环境筛的影响而很难发育成成熟个体[22],而较大年龄实生苗数量是保证种群持续发展的关键。在陇南山地,不同生境的5年生黄瑞香大龄苗数量均较低,除了受种群自身的生物学特性和生境异质性影响外,也不排除人为采挖大龄黄瑞香植株和幼苗被牛羊踩踏致死的可能。样地踏查时发现,大龄黄瑞香存在残桩萌枝的现象,这是黄瑞香应对不利环境的一种适应策略,也是维持种群繁衍和稳定的一种有效途径[23]。
生物量生产是能量流转于生态系统的结果,狭义生物量通常用干重表示,为特定面积或体积内所内含的生物物质质量,林木生物量分配的研究是分析林木结构功能的有效工具[24]。根冠比是群落中植物生物量存储方式和分配策略的重要指标[25-26],不同影响因子会引起植物地上、地下干物质分配比例发生变化,而植物也可通过改变各个器官生物量的分配和构型来调整对资源的获取方式[27]。黄瑞香实生苗根冠比半阳坡相对半阴坡较大,尤其是1年生幼苗的根冠比最大,这说明幼苗在半阳坡能更好的利用光照和温度等环境条件,故而能集中相对更多的资源至根部,储存更多的养分和水分,以备用于植株的营养和生殖[28]。半阳坡生境下,黄瑞香实生苗的茎较粗、植株高、分枝数多、根幅宽,说明较好的光照条件利于黄瑞香实生苗提高光竞争能力,形成壮苗。半阴坡生境下,黄瑞香实生苗总体看来植株细小、枝条叶片稀疏、根细而长,但单个叶面积较大,这说明实生苗在光热资源较差的环境中,植物会通过增加植株叶片面积和发展根系来增加对光照的吸收和能量的吸收,以期提高光竞争能力并获取更多的生长资源,这也是植物应对环境变化的一种生存策略[29]。本研究与胡晓静等[19]和余碧云等[27]对栓皮栎(Quercusvariabilis)的研究结果相似。
黄瑞香实生苗更新是环境因子和生境因子共同作用的结果,相关性分析结果显示,总盖度与黄瑞香实生苗高度和冠幅呈极显著负相关关系,灌木层盖度与黄瑞香实生苗冠幅呈显著负相关关系,说明黄瑞香野生群落所处的环境若是光照不足则不利于实生苗的生长壮大;土壤有机质、速效氮、速效磷和速效钾含量与黄瑞香实生苗冠幅、高度和根茎干重呈正相关关系,尤其是土壤速效氮,说明含有丰富氮元素的肥沃土壤更适合黄瑞香实生苗的生长发育。
4 结论
总之,在自然状态下黄瑞香天然更新能力不足、幼苗转化率低是制约其实生苗天然更新的关键环节,土壤条件和总盖度是影响实生苗更新的主要环境因子和生境因子,在今后陇南山地黄瑞香人工林经营中应尽可能的选择半阳坡生境。基于本研究结果,建议对黄瑞香天然更新地进行围封,降低人为挖掘大龄苗的概率和牲畜对幼苗的践踏伤害,而野生群落的更新抚育应通过改善土壤条件、疏枝除冠等措施改善实生苗更新环境,提高幼苗转化率,达到促进黄瑞香自然更新的目的。本研究将为黄瑞香野生种群的稳定更新和祖师麻药材的可持续利用奠定基础。
