新疆野苹果林野苹果实生苗空间分布及其生存现状
——以新源县为例
2018-05-07刘忠权董合干
刘忠权,董合干
1.伊犁职业技术学院,新疆伊犁 835000
2.伊犁州农村能源环境工作站,新疆伊犁 835000
新疆野苹果林是我国最大的苹果野生种自然分布群落[1],其中以新疆新源县野苹果林分布面积最大、密度最高[2~3]。在新源县野苹果林中,新疆野苹果(Malus sieversi)约占野果林的90%左右[4],由于长期以异交为主进行繁殖[5],故新疆野苹果种下变异类型极为丰富[6],存在诸多抗旱、抗寒、抗病虫、耐瘠薄类型[7],且该物种已被列为中国优先保护物种名录[8]。另有研究还表明新疆野苹果是栽培苹果的祖先种[9~10]。所以野苹果林是新疆非常重要的林业资源,对于新疆生物多样性保护、果树新品种选育及区域生态旅游均具重大意义。
近年来,由于人为干扰、过度采摘利用、过度放牧以及苹果小吉丁虫的危害等因素,新疆野苹果的自然种群数量和分布面积急剧减少,种群严重衰退[11~13]。实生苗更新是以种子为繁殖体所进行的有性生殖过程,是植物生活史中最重要的阶段[14],对促进种群进化维持种群遗传多样性意义重大[15]。实生苗作为种群更新的一种方式,由于其对不同环境的适应能力方面存在明显优越性[16],能提高或维持种群的遗传多样性不仅是野苹果种群天然更替的重要基础,也是实现野苹果种群可持续发展的前提条件。但关于新疆野苹果的研究多集中在资源调查[7]、果实形态与解剖[17]、遗传多样性[18]、种子萌发[19]、土壤种子库[20]、群落结构[21]和病虫害防治[22]等方面,关于新疆野苹果实生苗的相关研究未见报道。本研究将以新源县野果林新疆野苹果实生苗为调查对象,分析野苹果实生苗与海拔及其母株之间的空间关系,并对野苹果实生苗在野外环境下的生存状况进行观测,以期为新疆野苹果的保育和利用自然恢复提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
试验区地处伊犁河谷东部,气候湿润,冬暖夏凉;年平均气温7.7℃,1月平均气温~8.1℃,7月平均气温20.4℃,年降水量580 mm,≥10℃有效积温3006℃,日照时数2 674.5 h,土壤为山地黑棕色土,有机质含量12%~14%。
试验区内新疆野苹果主要在草原带以上和针叶林带以下的山地阴坡,其分布下限海拔约为1 000 m,上限海拔约为1 650 m,垂直分布的最适海拔在1 200m~1 400 m 之间,在此海拔之间可见野苹果纯林。在海拔1 200 m 以下常伴生有野杏(Armeniaca vulgaris)、野生樱桃李(Prunus cerasifera)、天山桦(Betula tianschanica Ruprecht)、准噶尔山楂(Crataegus songorica)等树种,海拔1 400 m 以上野苹果多零星分布,密度较小。
1.2 材料与方法
1.2.1 实生苗空间分布调查
由于野苹果分布下限受农业干扰较为严重,为减少农业活动对调查结果的影响,本次调查在海拔1300 m 以下不设置样方。调查于2014~2017年每年5月进行,根据地形以及新疆野苹果树的存活状况分别在海拔为1 366 m、1 439 m、1 489 m、1 600 m 处设置10 m×10 m 的样地。在样地内随机选择5 株野苹果树进行调查,并记录每株野苹果树的成活度、胸径、枝下高等数据;随后将该植株作为母株,并以母株为中心在其东南西北4个方向设置1 m×1 m 的样方,共计240个小样方,样方被精确到分成0.1 m×0.1 m 的正方形小格,分别调查距离母株0 m~1 m、1 m~2 m、2 m~3 m 之间实生苗的数量,并记录每个实生苗的坐标。调查中所有实生苗均为当年生幼苗,且在 2015年、2016年、2017年均未发现有实生苗。因此实生苗空间分布以2014年数据进行分析。
1.2.2 实生苗生存状况调查
在上述四个样地中,沿样地对角线上选择有幼苗的地方设置1 m×1 m 的小样方,每个样地设置5个共计20个小样方。每个小样方四角用木桩标记,同时用GPS 记录好坐标,小样方内的每一株实生苗的基部都用塑料标牌标记。调查时记录样方中当年生野苹果实生苗的数量、高度、健康状况、被啃食状况以及样方内草本植物的总盖度和平均高度。于2014年8月26日再次对上述标记的小样方进行调查。
1.3 数据分析
利用不同海拔样地内的240个小样方进行单因素方差分析,并用LSD 法对不同海拔实生苗数量进行多重比较。以20 棵野苹果树为研究对象分析比较距母株不同距离野苹果实生苗数量上的差异,并用LSD 发进行多重比较。数据分析采用spss19.0,作图使用origin9.0。
2 结果与分析
2.1 海拔与实生苗分布
由图1可知,总体上实生苗的密度虽然很低,但在不同海拔之间实生苗数量仍存在一定差异,表现为随着海拔的升高实生苗数量先增加后减小呈单峰型变化。从低海拔到高海拔实生苗数量分别为 0.38±0.16 株/m2、0.75±0.28 株/m2、0.3±0.19 株/m2、0.1±0.07 株/m2,方差分析结果表明除海拔1 439 m 与1 600 m 处实生苗数量分布差异达到显著水平(P<0.05)外,其余各海拔之间实生苗数量差异不显著。
在全部的240个小样方中只有28个小样方发现有实生苗存在,样方中出现实生苗的概率仅为11.6%。在28个样方中共调查到92 株实生苗,不同海拔实生苗数量占总实生苗数量分别为25.8%、48.4%、19.4%、6.5%(图1)。由数据可知海拔1 489 m 以下分布了约93.5%的实生苗,海拔1 439 m 处实生苗数量分布最多约占本次调查的50%左右;海拔为1 600 m 时,实生苗数量急剧下降,仅占总实生苗数量的6.5%。
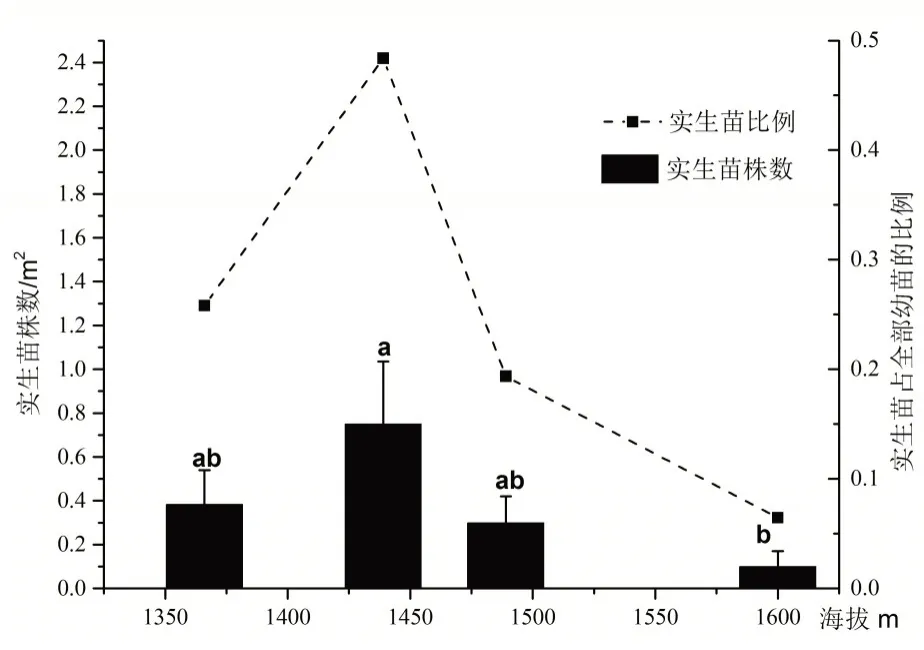
图1 海拔与实生苗分布
2.2 实生苗与母株距离的空间分布
以0.5 m 为单位分析实生苗与母株之间的距离(图2)。结果表明在0~0.5 m 范围内分布的实生苗数量占到总实生苗数量的7.6%;0.5~1.0 m 范围内分布的实生苗数量占到总实生苗数量的52.2%;1.0~1.5 m 范围内分布的实生苗数量占到总实生苗数量的17.4%;1.5~2.0 m 范围内分布的实生苗数量占到总实生苗数量的7.6%;2.0~2.5 m 范围内分布的实生苗数量占到总实生苗数量的2.2%;2.5~3.0 m 范围内分布的实生苗数量占到总实生苗数量的13%(图2)。总体表现为69.6%的实生苗集中在0.5~1.5这个范围内。

图2 实生苗与母株距离
距离母株距离的不同实生苗密度也有所变化,表现为0.5~1 m 处实生苗密度相对较高,可达 0.6±0.12 株/m2,而在 1.5~2.0 m 时实生苗密度相最低,仅为0.03±0.02 株/m2。(图2)对距离母株不同距离实生苗密度的方差分析结果显示,0.5~1 m 处的实生苗密度与其他距离密度差异达到极显著水平(P<0.01),其余距离的实生苗密度差异不显著。
2.3 实生苗的生存现状

表1 不同海拔实生苗的生存现状
在本次调查的20个固定小样方中,共跟踪了20 株幼苗的存活情况,在8月份的调查中全部死亡,其中11 株实生苗干枯死亡,6 株不见踪迹,3 株枯死但有被啃食痕迹。可见现阶段野苹果林中野苹果实生苗数量不仅少,而且这些实生苗无法存活到当年秋天,在自然状况下利用种子萌发的实生苗来更新野苹果种群非常困难。
由表1可知这20 株幼苗,从草本植物的盖度和高度上看,5月草本盖度从低海拔到高海拔依次为 46.6±5.7%、49.8±4.8%、50.4±5.1%、48±5.8%;到8月时盖度明显增加依次为85±2.2%、90±1.52%、88±3.32%、90±1.52%;草本植物的高度变化则更为明显,在5月草本高度从海拔到高海拔分别为4.8±0.3 cm、5±0.3 cm、4.4±0.5 cm、4.4±0.2 cm;到了 9月高度依次变为 59.8±9.8 cm、82.8±6.5 cm、91.8±7.6 cm、54.6±8.9 cm。这表明草本植物的盖度和高度对实生苗生长存在巨大影响,当年生野苹果实生苗生长在生长速度上不及草本植物,致使实生苗在光照和水分竞争中处于劣势而不能存活。
3 讨论
海拔高度的变化会影响到气候、水热等物理条件,致使不同海拔高度林下植被的变化,此种变化也使得幼苗分布在空间上发生变化,但不同地区不同物种的幼苗对海拔梯度变化的响应各不相同。研究表明坡垒(Hopea hainanensis)的幼苗数量和高度会随海拔的增加呈先下降后上升的趋势[23],而对天山北坡天山云杉天然更新幼苗的调查结果则表现为中部海拔最适宜幼苗的生长[24]。康冰[25]等对秦岭山地锐齿栎次生林幼苗研究认为,随着海拔的增高,土壤有机质及水分含量增加,为幼树提供了较为适宜的生长环境,而当海拔增加到临界高度,过低的温度成为了幼树生长发育的限制瓶颈,因此当海拔从1 083 m 增加到1 882 m 时,幼树密度变化有一个先增后减的趋势(图1)。本次研究结果与康冰等的结果类似,随着海拔的升高实生苗数量先增加后减小呈单峰型变化。可见海拔过高温度随之下降,较低的温度使得种子不能适时萌发最终失去活力或者种子萌发过程中更易遭受冻害而死亡。但这种环境条件下或许更容易孕育出高抗寒性的植株,因此,高海拔处的实生苗数量虽然较少,却对种群内部多样性的维持以及种群向外扩张具有重要意义。而低海拔地区,由于人为干扰相对较高(游客、放牧、采伐,采果等),实生苗的数量也较少。
株距同种子存活之间的空间联系的研究认为动物取食、病原体感染、幼苗竞争或种子取食者受距离制约影响。一般认为远离母株的繁殖体(种子、幼苗和幼树)与附近的繁殖体相比较,其生存机会较大,因此定居应该出现在远离母株的生境[26~27]。本研究结果显示野苹果实生苗绝大多数集中在母株冠幅内部,其中69.6%的实生苗集中在距母株0.5~1.5 m 范围内,13%集中在距母株2.5~3.0 m 范围内(图2)。其主要原因是人们为获得野苹果种子(用以做苹果嫁接的砧木)过度采摘果实。果实采摘通常会在8月中上旬结束,此时果实还未自然落地,因此采摘时均为机械手法,势必造成中部果实不易被采摘,待其完全成熟后自然落地而后进入土壤种子库,因此实生苗多集中在距母株0.5~1.5 m 处;而距母株2.5~3.0 m 则多野苹果树冠边缘,此处正是采果者收集果实的范围,一些个体小、有严重机械损伤的可能被遗弃从而进入土壤种子库获得成苗机会。但在该范围下实生苗均母株冠幅之下,光照相对较弱,不利于实生苗后期的生长,因此实生苗在种群自然更新中的作用十分有限。
实生苗对不同环境的适应能力方面存在优越性,能提高或维持种群的遗传多样性,对种群进化具有十分重要的意义[16]。但由种子转化成实生苗,是种群最易受外界环境影响、最敏感、最脆弱、面临死亡风险最高的阶段[28]。而木本植物的幼苗又更容易遭受干旱胁迫,其中幼龄幼苗由于没有足够的时间发育出横向伸展范围广、纵向延伸范围深的根系,可能最为敏感[29]。另外,林下的灌草层的生态学滤筛作用也是幼苗存活的潜在障碍,如Schiller 发现地中海松(Pinus halepensis)的幼苗在林冠覆被下的存活不会超过两年[30],此林冠下幼苗要想长大成为成年个体,就必须突破林下的灌草层。突破灌草层的幼苗会有较好的光照条件,从而为幼苗生长提供更多的光合产物,促进幼苗的高生长[31]。此外幼苗存活还受气温和降水的影响,高温加速土壤水分散失,形成干旱,抑制了幼苗的生长,而充足的降水可以保护幼苗不致旱死[32]。通过对四个样地,20 株实生苗的跟踪调查看,野苹果实生苗不仅数量少,更为严重的是当年生实生苗的存活率为0。野苹果林中实生苗数量少,一方面是由于人为过度开发利用及苹果小吉丁虫的危害致使结果数量显著减少[12];另一方面是野苹果果实较大,不易被地表的枯枝落叶和土壤埋藏,而且含储存物质多,因此更易遭受小哺乳动物和鸟类选择性地取食,从而影响了幼苗的数量。到8月实生苗存活率为0,主要是林下草本的盖度和高度显著增加,其中草本的盖度均在85%以上,高度均在60 cm 以上部分可达 100 cm 以上(表1),这些草本植物的存在与快速发展不仅影响了幼苗对地下营养与水的吸收而且草本根系使土壤紧密,影响幼苗的扎根[33],致使50%左右的实生苗在8月受水分胁迫干枯死亡。另有部分被啃食后死亡,部分幼苗丢失,说明野苹果实生苗处遭受水分胁迫外还常被其他动物取食。
通过上述分析可以发现研究区野苹果种群存在自然更新困难的问题。对实生苗存状况分析表明现阶段野苹果实生苗数量少且无法存活至当年秋季,可以预测在长期无新增个体补充且伴随苹果小吉丁虫危害的状况下,野苹果种群可持续发展问题十分棘手。针对现状,要解决野苹果种群的存活和发展问题,应该须采取综合性措施来抑制其种群的衰败趋势。首先,要及时建立自然保护区采取就地保护;其次,严厉禁止乱砍乱伐和采摘行为,努力保存现有种群个体的数量;再次,是大力发展人工辅助繁殖技术,探索幼苗最佳的移植年龄、寻找幼苗生长阶段的关键补水时段。因此,为了有效保护这一珍稀濒危树种资源,有关其幼苗生长与存活方面的研究仍需做大量工作。
4 结论
(1)不同海拔之间实生苗数量存在一定差异,表现为随着海拔的升高实生苗数量先增加后减小呈单峰型变化。
(2)距离母株距离的不同实生苗密度也有所变化,在0.5~1 m 处实生苗密度相对较高,且该处实生苗密度与其他距离密度差异达到极显著水平。但在该范围下实生苗均母株冠幅之下,光照相对较弱,不利于实生苗后期的生长,实生苗的这种分布限制了实生苗长成成年野苹果树的能力。
(3)野苹果幼苗数量极少,加上灌草层生长迅速,盖度较大实生苗均不能存活到当年秋天,同时实生苗在生长过程中易遭受水分胁迫和动物取食,说明在自然状况下,通过依靠种子萌发的实生苗来更新野苹果种群非常困难,因此必须采取人工辅助措施才能促进种群更新。
