荒漠植物柽柳抗逆机制的研究进展
2021-06-23李彩霞兰海燕
李彩霞 兰海燕
(新疆大学生命科学与技术学院 新疆生物资源基因工程重点实验室,乌鲁木齐 830046)
柽柳为柽柳科(Tamaricaceae)柽柳属(Tamarix Linn.)多年生灌木或小乔木,全世界分布约90种,中国有18种2个变种,主要分布于西北干旱、半干旱及华北滨海盐碱地区[1]。为了适应荒漠极端环境,柽柳在长期进化过程中形成了耐干旱、耐贫瘠、抗风沙等优良特性[2],并完善了泌盐系统,从而有效将盐离子从茎叶表面排出体外[3]。干旱区强烈的蒸发通常导致土壤表层盐分的大量累积,而柽柳主要利用地表水分和浅层地下水,因此,柽柳的生长常受到干旱和盐分双重胁迫[4]。逆境可导致植物细胞质渗透势升高、净光合速率(net photosynthetic rate,Pn)下降、二氧化碳(carbondioxide,CO2)利用率降低等[5],柽柳往往通过活性氧(reactive oxygen species,ROS)清除、代谢调节、蛋白质合成与再生等途径抵御胁迫伤害[6]。本文基于前人对荒漠旱生植物柽柳适应极端环境特性的研究,从柽柳种子形态及萌发特性、植株形态、生理及分子水平对其抗逆机制进行综述,旨为柽柳的深入研究提供理论依据。
1 柽柳种子形态特征和萌发休眠特性对逆境的适应
1.1 柽柳种子的形态特征及寿命
柽柳种子多为棒状或枣核状,种子顶端具芒柱,芒柱基部具长柔毛。成熟后的种子为深褐或浅褐色,毛呈浅褐或白色,种子极小,可借助风力进行远距离传播[7](图1)。柽柳种子大小与植株耐旱性相关,通过对12种柽柳耐旱性比较结果显示,沙生柽柳(Tamarix taklamakanensis M.T.Liu)耐旱性极强,其种子千粒重达410 mg,而其他柽柳种子千粒重则小于100 mg[8-9]。柽柳种子没有休眠现象,种子活力随贮藏时间的延长而迅速下降[10]。不同种类的柽柳种子寿命明显不同,如室温下刚毛柽柳(Tamarix hispida Willd)寿命为 11个月[11],多花柽柳(Tamarix hohenackeri Bunge)为 8 个月[12],甘蒙柽柳(Tamarix austromongolica Nakai)仅为 15 d[13]。

图1 多枝柽柳种子形态Fig.1 Seed morphology of Tamarix ramosissma
1.2 柽柳种子的萌发特性
在植物的整个生长周期中,种子萌发期是极脆弱的时期[14]。柽柳种群的不断繁衍得益于花期长、花量大、产种子时间跨度大等优势,并且种子通常可以借助水力和风力进行快速、广泛传播[15]。柽柳种子繁殖物候通常与河流流态特征相关,特别是洪水汛期和强度有着密切联系,由于柽柳具有双季开花结果的特征,其种子成熟期正好与夏季洪水泛滥期一致[16],此时,多枝柽柳种子可在河漫滩迅速萌发,由于根系的快速生长,使其在短时间内便可从地下吸收充足的水分以供地上部分生长发育,此后即使洪水改道,不再有地上水源供应,仍然可以利用地下水满足生存之需[17]。温度是种子萌发所需的另外一个重要条件,研究发现,多花柽柳可在5℃/15℃、5℃/25℃、15℃/25℃、25℃/35℃ 等 4个温变周期内萌发,较宽温度范围可使柽柳在不同季节利用水源进行萌发[18]。随着大气温度的升高,土壤表层水分不断蒸发,导致盐分的显著积累[19],从而抑制柽柳种子的萌发。当NaCl浓度高于100 mmol/L或聚乙二醇(Polyethylene glycol,PEG-6000)浓度大于30%(-1.2 MPa水势)时可显著抑制柽柳种子萌发,而50 mmol/L的NaCl则可促进中国柽柳种子的萌发[20-21],说明柽柳种子对盐分和水分胁迫有一定的耐受力。然而不同时期产生的柽柳种子耐受胁迫的程度不同,研究发现,虽然密花柽柳(Tamarix arceuthoides Bunge)春花期种子与夏花期种子的形状、大小、颜色、种子寿命均无显著差异,但春花期种子萌发速率高于夏花期[22],而夏花期种子耐盐、耐旱能力则高于春花期种子,这可能由于自然环境中夏花期种子面临的盐、旱胁迫远高于春花期而产生的一种适应性进化所导致[23]。
2 柽柳植株形态、结构特征及对逆境的适应
2.1 柽柳的叶片抗逆结构
叶片是植物进行同化作用并积累有机物和能量的主要器官[24]。柽柳叶片由表皮、皮层、维管柱三部分组成,表皮上具有下陷的气孔和盐腺,表皮细胞形状不规则且排列紧密,近轴面表皮细胞较叶肉细胞小,而远轴面则相反;表皮细胞向外突起形成高低不等的乳状突(图2-A),外面覆盖有角质层,不仅可以有效避免水分蒸发,而且有助于反射部分太阳光强辐射[25-26]。柽柳叶片上下表皮内侧均分布有栅栏组织,下表皮比上表皮发达,并均含有大量的叶绿体;栅栏组织与维管组织之间含有大量的薄壁细胞,有利于水分的贮存[27-28]。维管柱由维管组织和髓组成,维管组织为外韧型,其中初生韧皮部由厚壁细胞和薄壁细胞组成,次生韧皮部由8-12层细胞组成,排列紧密,髓部中央为大型薄壁细胞,可以保护维管组织免受短期水分胁迫[29-30]。
叶片肉质化、退化、硬化是荒漠植物适应极端环境的重要特征[31]。柽柳幼苗期叶片肉质化,成株叶片叶柄退化,抱茎而生,形成茎-叶复合体,大大减小了暴露在空气中的叶面积(图2-B和图2-C);不仅如此,由于互生叶密集排列,在减少水分散失的同时还增加了营养枝的有效光合面积[32]。柽柳叶片对逆境具有较强的适应能力,干旱胁迫促使其迅速完善旱生结构,如栅栏组织增加、角质层增厚、表皮细胞面积减小等,并能通过调节叶片的抱茎程度,进而适应不同逆境的胁迫[33]。此外,柽柳叶皮层中的薄壁组织还具有较强的可塑性,可发育成不同特化程度的厚壁组织,在增加叶片机械支撑力的同时还能防止水分的蒸腾损失[34]。
2.2 柽柳根系结构对逆境的适应

图2 不同时期多枝柽柳叶片形态Fig.2 Leaf morphology of Tamarix ramosissma in different developmental stages
荒漠植物应对极端环境除了地上部结构如叶片、气孔、盐腺等的调节外,其地下根系的适应性变化是另一种适应对策[35](图3)。柽柳是一种兼性潜水植物,根系极其发达,主根可深入至地下水层。当地下水位较浅时,柽柳根系可直接吸收地下水,而当地下水位降至根区以下则可从周围土壤中吸取水分,因而对干旱有很强的耐受性[36-37]。在降雨较多的季节其根系主要利用土壤速变层的水分,而降雨较少时则利用活跃层及相对稳定层的水分[38]。柽柳的细根(直径小于2 mm)大量分布于40-60 cm的土层中,土壤水分的快速蒸发可促进柽柳水平根分生出垂直方向的深根[39],因此,柽柳的根系可以利用多层土壤中的水分,包括对降雨和深层地下水的吸收,并可将深土层中的水分及地下水输送并释放到干土层中,这种由水势梯度驱动的现象被称为“水力提升(hydraulic lifting)”[40]。由于白天柽柳地上部分的蒸腾作用导致茎中的水势小于土壤水势,植物正常吸水供给生长;而18:00后蒸腾作用减弱,当主根吸水完成组织失水的再填充后再供给主枝用以夜间蒸腾,导致茎中水势逐渐升高,在21:00后茎水势高于土壤水势,由于根水势恢复速度快于茎水势,且根系水势大于茎水势,因而形成根系水势大于浅层土壤水势,深层土壤的水分因此能从植物根系释放到浅层干燥的土壤层中,这为柽柳“释水”现象提供了解释[41]。基于此,柽柳地上部分强烈蒸腾作用引起的生理干旱得以缓解,柽柳这种在不同时间段利用不同土壤层水分的能力,可能是其耐受逆境并保证养分吸收的一个重要原因[42]。此外,多枝柽柳还可在靠近地下水位的根际萌生新根,从而快速补充地上植株高温蒸腾导致的水分损失,由此呈现出的柽柳根系结构及水分在不同层级根系间的运输调配,可能是柽柳应对荒漠干旱及高温失水的重要策略[43]。
植物对水分、养分的吸收与根系的侧根密度、构型及生物量密切相关。柽柳根系结构随环境及发育阶段呈现不同的类型,一年生植株的根系呈鱼尾形,四年生则呈叉状,这是适应生境的一种体现[44]。多枝柽柳根系密度、生长量与土壤盐分含量成反比,显示盐分限制侧根的发生和分布[45]。随着盐浓度的升高,柽柳根冠比明显增加,暗示柽柳根冠间存在着异速生长关系,并通过这种生长调节来适应逆境[46]。

图3 柽柳植株地上地下协同吸水、保水调控机制(参考文献[47-49]绘制)Fig.3 Schematic diagram of water absorption and retention regulation of Tamarix above and under ground parts of plant (drawn by reference[47-49])
3 柽柳盐腺结构及泌盐机制
3.1 盐腺结构
植物为了适应生境往往产生趋同或趋异进化现象,外部形态上的表现尤为明显[50]。盐腺是柽柳适应盐碱环境的一个重要形态特征,主要分布于茎和叶上,可将细胞内多余盐分有效排出体外[51]。盐腺可分为双细胞盐腺和多细胞盐腺,柽柳是多细胞盐腺的典型代表[52]。柽柳盐腺由8个细胞组成,内层2个收集细胞,外层6个分泌细胞,收集细胞具有大液泡而分泌细胞具有浓缩的细胞质,盐腺外部被厚厚的角质层包裹[53]。当腺体完全形成并泌盐时,分泌细胞的细胞壁局部向细胞质延伸形成棒状突起,说明突起与分泌过程直接相关。分泌细胞与收集细胞被表皮层包围,这种非角质化的区域称为融合区,该区内的分泌细胞与收集细胞由胞间连丝连接[54]。胞间连丝是盐分进入腺体的主要途径,并且盐分由一个细胞移动到另一个细胞也是借助胞间连丝完成的[55](图4-A 和图4-B)。
3.2 盐腺泌盐机制
盐腺按泌盐方式可分为2种:一种是直接向叶表面分泌盐(向外泌盐的盐生植物);另一种是将盐收集在特定液泡中(向内泌盐的盐生植物)[56]。柽柳盐腺泌盐方式属于前者,且优先选择分泌二价阳离子,其次是一价阳离子,而Na+是一价盐离子的首选[57]。离子在盐腺细胞间和盐腺与叶肉细胞间的运输主要通过质外体途径和共质体途径来完成,前者是离子从盐腺基部无角质层包裹的区域进入盐腺,然后通过分泌孔将盐离子分泌到体外;后者是离子在盐腺各组成细胞之间进行转运[58]。研究发现,无叶柽柳[Tamarix aphylla(L.)H.Karst.]将盐离子排出体外后,盐腺会逐渐衰老并最终死亡,衰老的盐腺在幼叶和成熟叶中均存在,说明柽柳叶片上的盐腺结构存在新老更替的动态变化过程,一些盐腺完成使命后,新的盐腺会继续分化[59]。当盐腺进入衰老期后,首先破坏的是2个收集细胞,其次是6个分泌细胞,在此过程中,分泌细胞质中会形成很多自噬泡,导致细胞变性并吞噬和消化细胞质,随后核膜和内质网不断扩张,最终导致分泌细胞解体,进而被具有泌盐功能的新腺体所取代[60]。
3.3 影响盐腺泌盐的因素
柽柳可以分泌多种类型的离子,其中,二价阳离子有Mg2+和Ca2+,一价阳离子有Na+和K+,阴离子有SO42-和Cl-,在逆境下,Na+、Cl-分泌量最高,SO42-次之,K+、Mg2+和 Ca2+的分泌量较低[61]。盐腺分泌离子与多种因素相关,如昼夜节律、土壤盐离子含量、温度、湿度、土壤水分等(图4-C)。相关研究表明,柽柳盐腺的泌盐频率与大气湿度呈正相关,与大气温度呈负相关,与光照强度、蒸腾作用无明显相关性。通常盐离子的分泌上午高于下午,在8:00-10:00的分泌率最高;而日间则高于夜间[62-63]。随着盐、旱胁迫的增强,柽柳的盐腺密度和总泌盐速率均上升,但当NaCl浓度超过300 mmol/L时,盐腺密度下降;而在干旱胁迫下柽柳泌盐活动明显减弱,单个盐腺泌盐速率降低以防止水分散失[64]。
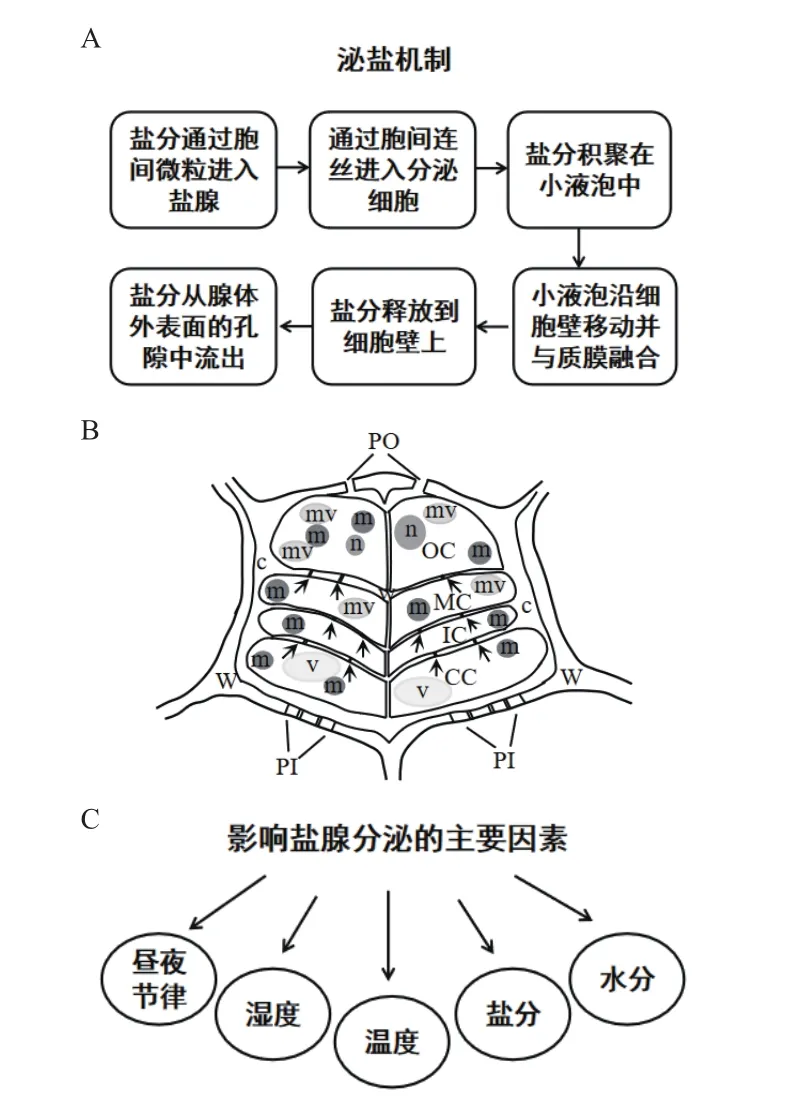
图4 柽柳的盐腺结构、泌盐机制及泌盐影响因素模式图(参考文献[55,65-66]绘制)Fig.4 Schematic diagram of salt gland structure, secretion mechanism and influencing factors in Tamarix (drawn by reference [55, 65-66])
4 柽柳抗逆的生理机制
4.1 渗透调节系统对逆境的响应
4.1.1 离子吸收 离子稳态的建立是植物耐盐的重要策略,柽柳通过将盐离子排出体外从而维持体内离子平衡[67]。随着盐分的积累,柽柳体内Na+、Cl-含量逐渐上升,K+、Ca2+和Mg2+则呈下降趋势,显示各种离子的吸收和积累呈现相互制衡的关系[68]。在同等盐分水平下,刚毛柽柳体内Na+和K+含量依次为叶>根>茎,且Na+含量始终大于K+,说明叶片是Na+和K+积累的主要部位,通过茎的传输,使其他部位不会出现盐离子的累积效应[69]。柽柳对离子的吸收与土壤中的离子分布和含量有关,通常主根周围土壤中阳离子富集程度依次为Na+>K+>Ca2+>Mg2+, 阴 离 子 为土层越深,Na+、Ca2+、Mg2+和Cl-含量越高,显现出底聚特征;而和HCO3-含量则随着土层深度而降低,呈表聚特征,这部分原因可能是柽柳浅层土壤中的侧根较多,吸收的阳离子多于深层土壤,而且雨水淋溶过程也会导致盐分从浅层不断迁移至深层[71]。
4.1.2 渗透调节物质 有机渗透调节物质如脯氨酸、可溶性糖、甜菜碱、可溶性蛋白质等,可通过维持膨压和特定的溶质从而调节植物细胞水势并维持细胞的正常功能[72-73]。游离脯氨酸、可溶性糖浓度的增加是水分胁迫的一个重要指标。相关研究表明,中国柽柳的叶、茎和根部脯氨酸含量随盐浓度的上升而增加;而甜菜碱、可溶性蛋白含量未见显著变化,由此显示,脯氨酸在柽柳应对高盐环境中发挥着重要的作用[74-75];然而在重度盐、旱胁迫下柽柳可溶性糖含量则呈现下降趋势[76]。除此之外,渗透调节物质的积累与柽柳的抗寒性也呈正相关,随温度下降,柽柳脯氨酸及可溶性糖含量均呈上升趋势[77]。上述研究表明,渗透调节物质在柽柳应对胁迫过程中发挥着重要作用。
4.2 抗氧化系统对逆境的响应
逆境可导致植物体内ROS的产生,随后攻击细胞中的大分子,产生DNA损伤、膜透性改变、蛋白质合成紊乱等[78]。细胞中的抗氧化酶则能清除ROS,研究发现,低盐浓度(≤100 mmol/L)下多枝柽柳过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)、超氧化歧化酶(superoxide dismutase,SOD)的活性升高,从而有效清除多余的ROS,使丙二醛(malondialdehyde,MDA)保持在较低水平;然而随着NaCl浓度的不断升高(≥200 mmol/L),SOD、POD活性减弱,导致ROS不断积累,膜脂被过氧化[79-80],膜透性增大,细胞内溶物外渗,最终导致植物组织被破坏[81]。柽柳体内抗氧化酶对逆境的反应速度依柽柳品种不同而有差异,在低温胁迫下(0℃以下),盐地柽柳(Tamarix karelinii Bunge)、密花柽柳(Tamarix arceuthoides Bunge)、长穗柽柳中的SOD活性急剧升高,而刚毛柽柳和翠枝柽柳(Tamarix gracilis Willd)则呈缓慢上升的趋势[82];在55℃高温条件下,多枝柽柳CAT活性迅速升高,60℃时又大幅下降,可能是高温破坏了酶蛋白结构进而导致活力下降。在植物体内,SOD、POD、CAT必须协调一致才能将ROS维持在较低水平从而避免对膜的伤害[83]。与此同时,抗氧化剂也可以清除一部分氧自由基。在干旱胁迫下,多枝柽柳不仅可以在体内合成多酚物质降低细胞渗透势,多酚还可以通过化学方式结合一部分自由水,进而缓解水分亏缺[84]。研究发现,柽柳叶片中的黄酮可清除羟自由基(·OH)及DPPH(1,1-二苯基-2-三硝基苯肼)自由基[85],黄酮类化合物柚皮苷可抑制 ABTS+(2,2′-联氮-双-3-乙基苯并噻唑啉-6-磺酸)自由基,作为抗氧化剂进一步提高了抗氧化系统的可靠性[86]。
4.3 光合作用系统的响应
光合作用是植物生长发育的主要驱动力。多枝柽柳Pn、光能利用率(light utilization efficiency,LUE)呈不对称双峰型日变化趋势,Pn和LUE主峰均出现在10:00,前者的次峰出现在14:00,后者在16:00;水分利用率(water utilization efficiency,WUE)在11:00和16:00分别有2个不明显的峰;而蒸腾速率(transpiration rate,Tr)日变化则为单峰型,峰值出现在12:00,由此显示多枝柽柳的Pn、LUE、WUE等在午间会出现光抑制现象[87-88]。研究表明,干旱季节刚毛柽柳的WUE上午显著高于下午,此时Tr下降幅度大于Pn,从而提高WUE[89];而中国柽柳为了降低日耗水量,采取推迟清晨茎部汁液流动开始的时间,并提前结束日间汁液流动,从而保证正常的生长代谢过程[90]。此外,随着光合有效辐射(photosynthetically active radiation,PAR)的增强,不同柽柳种在达到光饱和点后Pn并未出现明显下降,而Tr呈下降趋势,暗示柽柳可能通过降低蒸腾速率维持正常的光合生理需水[91]。
盐胁迫会导致叶绿体基粒和基质类囊体减少,进而影响光合效率[92]。中国柽柳在低盐胁迫下能够保持较高的Pn、Tr和树干液流速率[93],但随盐分的升高,这些指标及叶绿素a/b含量下降,类胡萝卜素/总叶绿素比值增加,显示高盐加剧柽柳光合色素分解,进而使光合效率降低。同时,胞间CO2浓度(intercellular CO2concentration,Ci)、气孔导度(stomatal conductance,Gs)呈先下降趋势,而气孔限制(stomatal limitation,Ls)值则相反[94-95],由此显示,适当的盐分可以激活柽柳光合作用;而高盐环境下,柽柳组织出现生理缺水并导致气孔孔径减小甚至关闭,改变气体交换频率和蒸腾作用,进而影响光合作用[96]。
5 柽柳响应逆境胁迫的分子调控机制
植物逆境应答信号传导主要由转录因子和信号级联系统中的各种激酶协同完成[97-98]。其中丝氨酸/苏氨酸激酶(serine/threonine kinase)、钙依赖蛋白 激 酶(calcium-dependent protein kinase,CDPK)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、G蛋白等在柽柳响应盐旱胁迫信号过程中具有重要作用[99]。相关研究表明,柽柳转录因子基因ThbHLH的启动子序列含有DOF、Ibox、W-box等与抗逆应答相关的顺式作用元件,将ThbHLH转入拟南芥,可以明显降低O2-、H2O2水平[100]。并且转录因子 ThCRF1可与 TTG、DRE、GCC等基序结合,诱导吡咯-5-羧酸合成酶(P5CS)、海藻糖-6-磷酸磷酸酶(TPP)、SOD和POD等基因的表达,从而提高脯氨酸、海藻糖的含量,并增强 POD 和 SOD 的活性[101]。过表达 ThbZIP1、ThWRKY4、ThMYB13等可增强刚毛柽柳对ROS的清除能力,促进渗透调节物质的积累和可溶性蛋白质的生物合成,维持K+/Na+平衡,进而提高柽柳的抗逆能力[102-104]。
近年来,已鉴定出许多柽柳逆境诱导基因的功能,如胚胎晚期发育丰富蛋白基因(LEA)、吡咯啉-5-羧酸还原酶基因(P5CR)、质膜内在蛋白基因(TIP)、翻译起始因子基因(eIF)、类锌指基因(ZFL)、冷适应蛋白基因(CAP)等。研究发现将柽柳ThLEA转入烟草可降低干旱胁迫下植株的膜脂过氧化程度,转入蓝莓可增强低温胁迫下植株的SOD和POD酶活性,显示LEA可通过细胞膜免受破坏来提高柽柳的抗性[105-106]。试验证明,在盐、旱胁迫下,过表达ThP5CR显著提高刚毛柽柳体内脯氨酸含量,增强细胞ROS的清除能力[107]。张春蕊[108]发现过表达柽柳水通道蛋白基因ThTIP,不仅可以提高盐、旱胁迫下拟南芥的抗氧化酶活性以及脯氨酸和叶绿素的含量,而且还能降低植物失水率和MDA含量。杨桂燕等[109]将ThGSTZ1转入酿酒酵母可以提高酵母对低温和高温的耐受性。可见这些逆境诱导基因可从渗透调节物质积累、抗氧化等多方面使柽柳适应短期的非生物胁迫。
6 展望
非生物胁迫严重影响植物生存,柽柳经过不断进化形成了多种适应性特征,包括形态结构、生理生化及分子生物学的不同层面。前期已对柽柳抗逆性开展了广泛的研究,但由于柽柳为多年生灌木,对野外自然生长植丛的研究比较困难,室内又难以长期模拟自然生境条件,加之苗期生理状态与成株存在较大差异,导致获得数据不能全面反映实际情况。因此,在今后的研究中,应着力改进室内或室外柽柳的培养条件,尽可能接近自然生境,促进柽柳抗逆机理研究的准确性与高效性。近年来,对柽柳抗逆相关基因的克隆促进了其相关作用机制的研究,但进展相对缓慢,亟待将组学、生物化学等多种手段结合应用于柽柳抗逆机理的研究,从而深入阐明柽柳抗逆相关机制,并为柽柳在生态环境改良及抗逆农作物选育方面的应用提供理论依据。
