黔西北地区云南松种群数量动态及空间分布格局
2021-06-07李仰征李望军
何 斌,李 青,李仰征,李望军
(1.贵州省典型高原湿地生态保护与修复重点实验室,贵州 毕节 551700;2.贵州工程应用技术学院 生态工程学院,贵州 毕节 551700)
0 引言
种群作为群落结构和功能的基本单位,具有数量特征、遗传特征、空间分布特征和系统特征等[1],其数量动态和空间分布格局一直是植物种群生态学的重要研究内容[2]。种群数量动态是种群与环境相互适应的结果,可以客观反映种群在时间尺度上的变动规律和演替趋势[3]。对种群数量动态进行研究,有利于人们全面地认识种群的形成机制、影响因素、更新策略和演替规律等[4],揭示种群与环境之间的关系,在种群的管理和生态系统的恢复方面具有重要的应用价值[5]。种群空间分布格局是物种生物学特性、种间关系和环境条件相互作用的结果[6]。种群空间分布格局不仅与物种自身生物学特性有关,而且会受到发育阶段和生境条件的直接或间接影响[7]。分析种群空间分布格局有助于认识种内、种间的相互作用及种群分布与环境的关系[8],揭示种群格局形成的原因,为生物多样性保护及物种保育提供科学依据[9]。因此,研究种群数量动态和空间分布格局对阐明种群的发展机制和各种生态过程及群落的稳定性和演替规律等具有重要意义[10]。
云南松(Pinusyunnanensis)是中国特有树种,主要分布在云南、贵州和四川等地区,具有耐干旱瘠薄、适应能力强等特点,是荒山造林的先锋乡土植物,具有重要的生态和经济效益。目前,国内学者从物种多样性、生物量、土壤养分和生态系统功能等方面对云南松进行了大量研究[11-13],但对云南松种群的数量动态和空间分布格局的研究报道较少[14-15],而且这些研究主要是针对云南省分布的种群进行的。已有研究表明,同一物种在不同地区、不同尺度下的种群结构和空间分布格局可能不同[16-18]。那么,云南松作为贵州省西北地区的一种重要森林资源,其年龄结构如何?群落究竟处于哪种状态? 空间分布格局是怎样的? 种群结构的维持机制是怎样的?
鉴于此,本文以贵州省威宁县境内的云南松林为研究对象,采用典型样地法进行野外调查,应用“空间代替时间”的方法编制种群静态生命表,分析种群的数量动态,采用Cassie 指数(Ca)、丛生指数(I)、扩散型指数(I&)等指标分析种群的空间分布格局,试图探讨以下问题:1)云南松种群的数量动态是怎样的;2)云南松种群的空间分布格局是怎样的;3)不同生长阶段云南松种群空间分布格局是否存在差异。通过本研究,认识贵州省西北地区云南松林种群数量动态和空间分布格局,以期为该地区云南松林的可持续经营提供理论基础,为喀斯特地区森林生态系统的管理和保护提供参考。
1 研究区概况
研究区位于贵州省毕节市威宁县(26°47′32″~26°52′52″N,104°10′16″~104°20′40″E),处于东南季风与西南季风的过渡带,属于亚热带季风性湿润气候区。年平均气温10.5 ℃,年平均降水量950.9 mm,主要集中在5—10月;光照充足,年平均日照时数1 800 h,无霜期180 d;地形以山地丘陵为主,平均海拔2 200 m;土壤类型以黄棕壤、石灰土和石质土为主。
该地区的云南松林乔木层主要有云南松、槲栎(Quercusaliena)和华山松(Pinusarmandii),灌木层主要有扁刺峨眉蔷薇(Rosaomeiensis)、金花小檗(Berberiswilsonae)、牛奶子(Elaeagnusumbellata)、矮杨梅(Myricanana)、火棘(Pyracanthafortuneana)、西南栒子(Cotoneasterfranchetii)、荚蒾(Viburnumdilatatum)、滇榛(Corylusyunnanensis),草本层主要有一年蓬(Erigeronannuus)、车前草(Plantagoasiatica)、紫花地丁(Violaphilippica)、鱼腥草(Houttuyniacordata)、云南莎草(Cyperusduclouxii)、夏枯草(Prunellavulgaris)、风轮草(Clinopodiumpolycephalum)等。
2 研究方法
2.1 样地调查
在对威宁县云南松林分布区全面踏查的基础上,2018年7—9月选择在集中连片、有代表性的区域设置20 m×30 m的样地12块。采用相邻格子法将每块样地划分成24个5 m×5 m 的小样方,采用每木检尺对每个小样方内的乔木进行调查,记录树高、胸径、冠幅和个体相对坐标(x,y)等,同时记录各样方内的灌木和草本植物种类、盖度和多度等,并记录各样地的经纬度、海拔、坡向等基本环境因子(表1)。
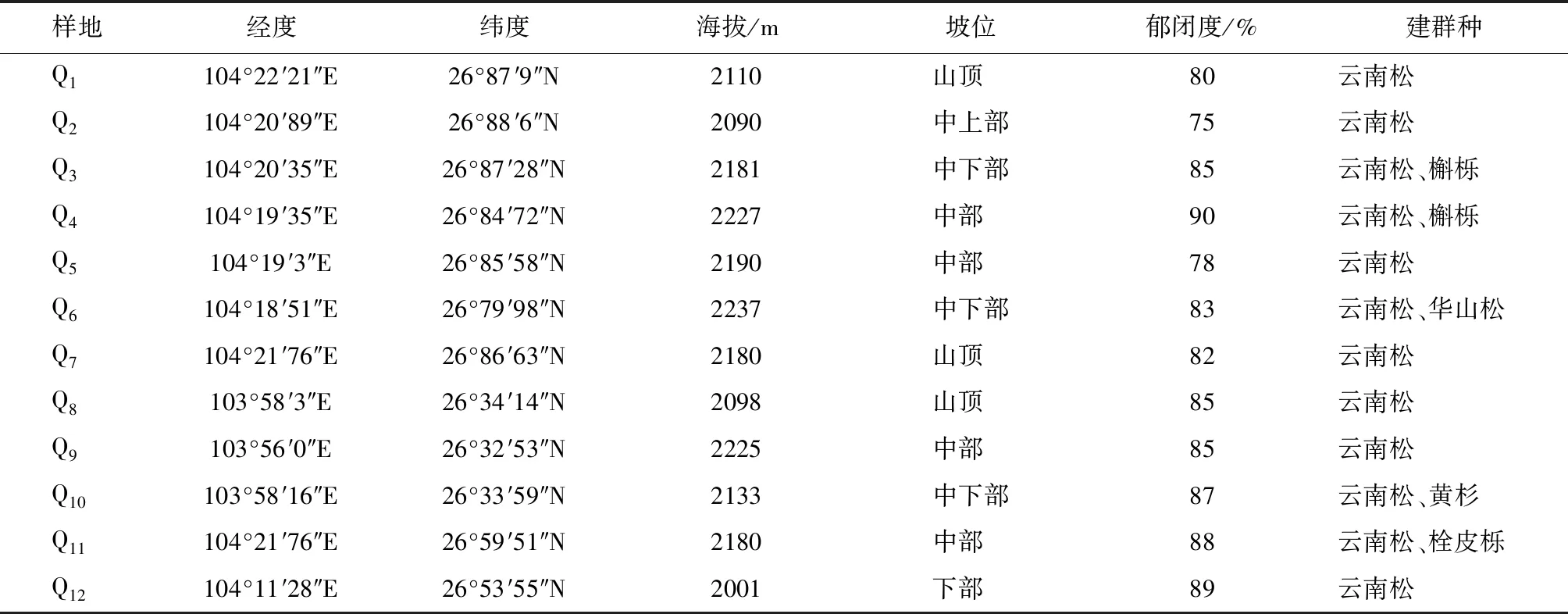
表1 样地信息Tab.1 Information of plots
2.2 龄级划分
在相同的环境条件下,同一树种径级和龄级的生长规律具有一致性[19]。因此,本文采用径级结构代替年龄结构来研究云南松的种群结构。根据云南松的调查数据,结合其生长发育特点,云南松种群被划分为9个等级(即9个龄级)。龄级Ⅰ:DBH<2.5 cm;龄级Ⅱ:2.5 cm≤DBH<5 cm;龄级Ⅲ:5 cm≤DBH<9 cm;龄级Ⅳ:9 cm≤DBH<13 cm;龄级Ⅴ:13 cm≤DBH<17 cm;龄级Ⅵ:17 cm≤DBH<21 cm;龄级Ⅶ:21 cm≤DBH<25 cm;龄级Ⅷ:25 cm≤DBH<29 cm;龄级Ⅸ:DBH≥29 cm。分别统计不同龄级的植株个体数,编制静态生命表。
2.3 静态生命表和存活曲线
静态生命表是根据某一特定时间种群不同龄级间的个体数描述种群生活状态的一种方法[20],主要包括ax(x龄级内的现存个体数)、lx(x龄级开始时标准化存活个体数)、dx(x到x+1龄级间隔期内的标准化死亡数)、qx(x到x+1龄级间隔期间的死亡率)、Lx(x到x+1龄级间隔期间还存活的个体数)、Tx(x龄级到超过x龄级的个体总数)、ex(进入x龄级的平均期望寿命)、Sx(存活率)、Kx(消失率)等相关参数,参考文献[4]中的计算公式。
由于静态生命表是根据实际调查数据由“空间推时间”和“横向导纵向”方法推导种群动态过程,而不是对某一种群全部生活史的追踪,不可避免地会产生与静态生命表编制要求的3个数学假设不符的现象,出现死亡率为负的情况。许多学者采用匀滑技术处理统计数据,本研究参考赵阳等[21]的方法,采用方程拟合法对数据进行匀滑处理。
存活曲线是描述种群个体在各龄级生存状况的一条曲线。根据Deevey[22]的理论,存活曲线分为3种类型,即Deevey-Ⅰ,Deevey-Ⅱ和Deevey-Ⅲ 3种类型。根据Hett等[23]的理论,用指数函数Nx=N0e-bx和幂函数Nx=N0x-b对云南松种群存活曲线进行检验。
2.4 种群动态量化分析
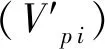
(1)
(2)
(3)
(4)

2.5 空间分布格局
测定种群空间分布格局的数学模型较多,不同模型有其不同的优势和不足。为了保证分析结果的有效性和合理性,本研究根据样地调查时林木的坐标(x,y),采用Caasier 模型中的扩散系数(C)、Lloyd 模型中的平均拥挤度(M*)和聚块性指数(PAI)、Davidh & Moore 模型中的丛生指数(I)、Morisita 模型中的扩散型指数(I&)和Kunou 模型中的Cassie 指数(Ca)来计算分析云南松种群及其不同发育阶段的分布格局。各指数的计算和评判标准参考文献[25]中的方法。
3 结果与分析
3.1 云南松种群年龄结构特征
由图1可知,云南松种群龄级结构呈纺锤型,个体主要集中在Ⅲ—Ⅵ龄级,占比高达78.93%,Ⅰ—Ⅱ龄级个体数占总数的11.87%,说明云南松种群中龄林个体数充足,但幼龄个体数占比较少,比较匮乏;Ⅶ—Ⅸ龄级个体数仅占总数9.20%,且呈阶梯式下降趋势。由此可见,云南松种群处于前期薄弱、中期稳定、后期衰退的状态。
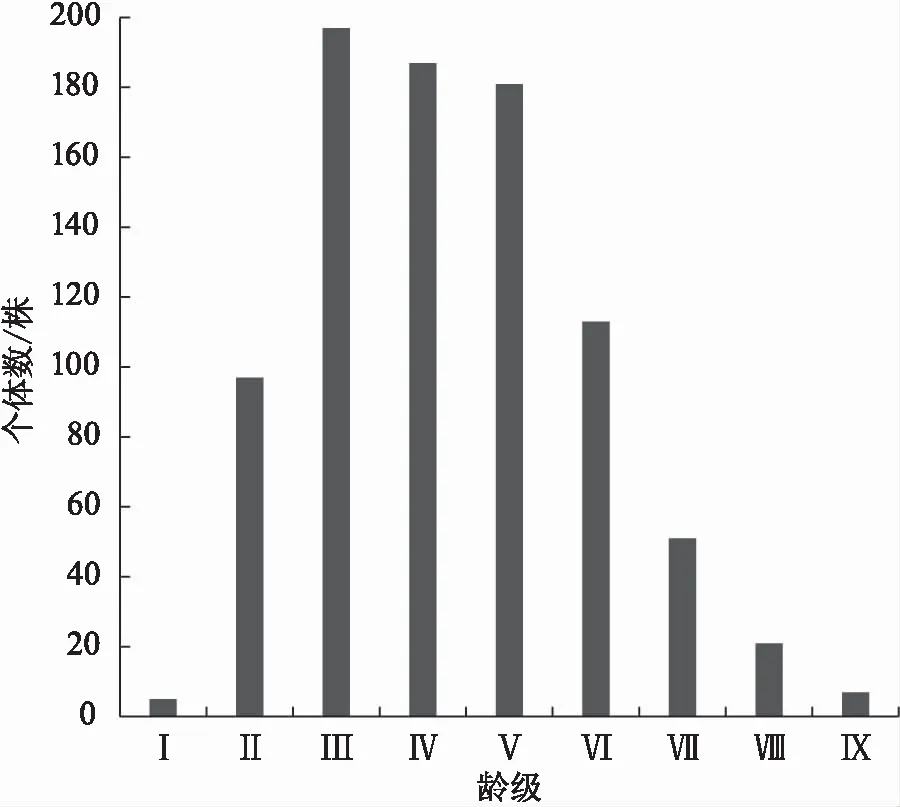
注:“Ⅰ”表示DBH<2.5 cm;“Ⅱ”表示2.5 cm≤DBH<5 cm;“Ⅲ”表示5 cm≤DBH<9 cm;“Ⅳ”表示9 cm≤DBH<13 cm;“Ⅴ”表示13 cm≤DBH<17 cm;“Ⅵ”表示17 cm≤DBH<21 cm;“Ⅶ”表示21 cm≤DBH<25 cm;“Ⅷ”表示25 cm≤DBH<29 cm;“Ⅸ”表示DBH≥29 cm。图1 云南松种群径级结构Fig.1 Diameter class structure of P.yunnanensis population
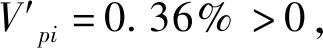

表2 云南松种群动态变化指数Tab.2 Dynamic change index of P.yunnanensis population
3.2 云南松种群静态生命表及存活曲线
云南松种群静态生命表(表3)表明,随着林龄的增加,生命期望值(ex)整体上呈下降趋势,Ⅰ龄级的生命期望值(ex)最高,为3.5,表明该阶段云南松的生存环境质量高,群落内的水、热和营养等条件丰富,个体间竞争小,能有效地利用资源;随着林龄的增加,个体生长所需的生长空间和营养条件逐渐增加,同时,郁闭度逐渐增加、光照条件不足,相互间竞争逐渐加剧,生命期望值(ex)逐渐下降,这与云南松的生物学特性一致。值得一提的是Ⅷ龄级的生命期望值(ex)有小幅回升,这可能是由于云南松个体在Ⅶ龄级的大量死亡,为其他个体的生长创造了有利的生长条件。
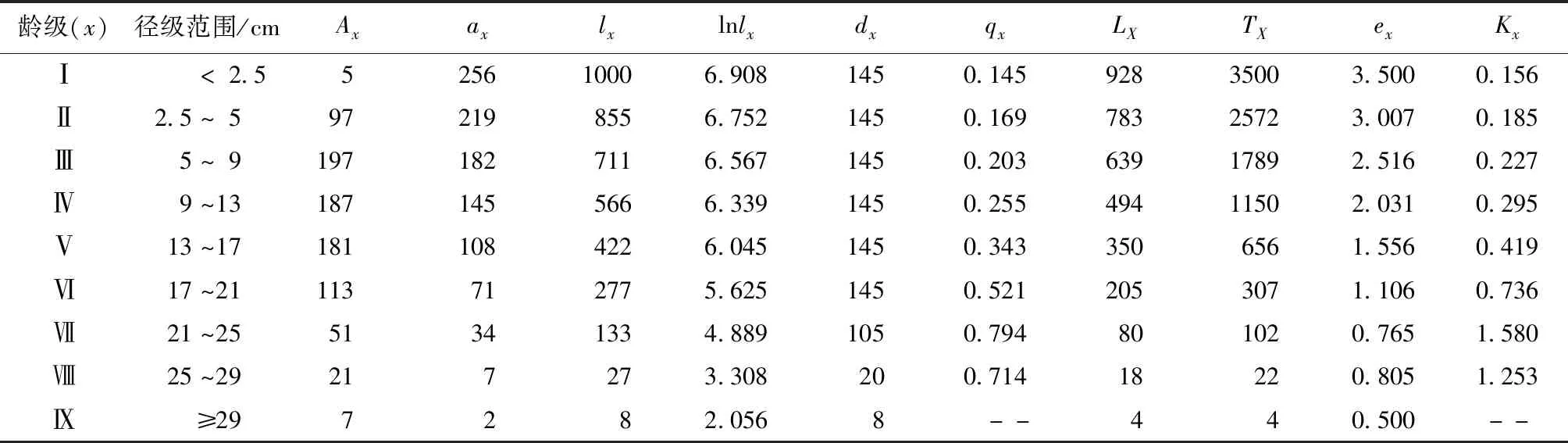
表3 云南松种群静态生命表Tab.3 The static life table of P.yunnanensis population
由表3可知,随着林龄的增加,云南松种群的死亡率和消失率总体上呈增加趋势,在Ⅶ龄级急剧上升,达到峰值,死亡率和消失率分别为0.794和1.580。说明此阶段云南松已经进入了生理衰退期。这可能是由于随着云南松个体的生长,植株个体间竞争加剧、生态位重叠,林内的光照、水分等营养条件和空间环境均无法满足其生长需求,自疏和它疏作用逐渐显现,个体间分化严重,导致大量个体死亡。
由图2可知,云南松种群存活率整体呈下降趋势,第Ⅰ—Ⅵ龄级存活曲线斜率较为平缓,第Ⅵ龄级后曲线斜率突然变大,云南松存活数下降较快,死亡个体较多。与Deevey[22]理论中的经典存活曲线比较,云南松种群存活曲线介于Deevy-II型和Deevy-III 型之间。应用Hett等[23]的数学模型检验云南松种群的存活曲线,建立如下模型:
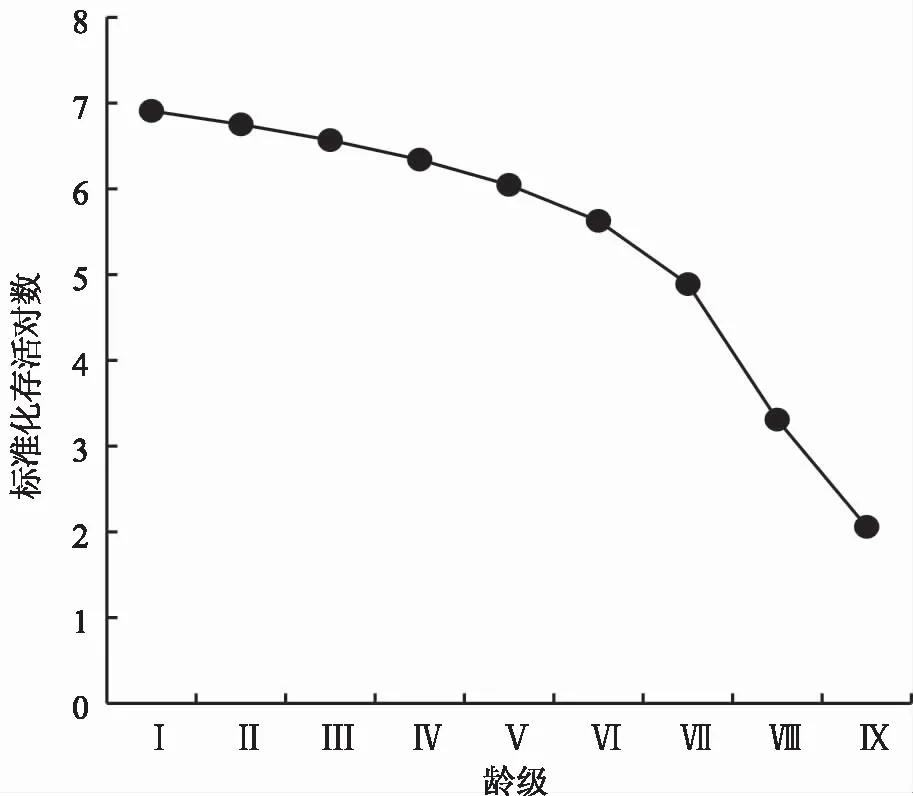
图2 云南松种群存活曲线Fig.2 The survival curve of P.yunnanensis population
Y=9.6203e-0.128xR2=0.7399
Y=8.9872x-0.403R2=0.5039
由于指数模型的R2(0.7399)显著大于幂函数模型的R2(0.5039),因此,云南松存活曲线更趋近于 Deevy-II 型。
3.3 种群空间分布格局
从云南松种群的空间分布格局(表4)中可以看出,扩散系数(C)、聚块性指数(PAI)、丛生指数(I)、扩散型指数(I&)和Cassie指数(Ca)的计算结果均表明云南松种群呈集群分布格局。不同样地云南松种群空间分布的各项指数差异较大,说明其集聚程度不同。从平均拥挤度指数(M*)来看,其它样地的M*指数明显小于样地Q1,Q9和Q12,同时,其它样地的扩散系数(C)和扩散型指数(I&)也明显小于这3块样地,这主要是由于Q1,Q9和Q12这3块样地中云南松立木株数较多、胸径较小造成的。
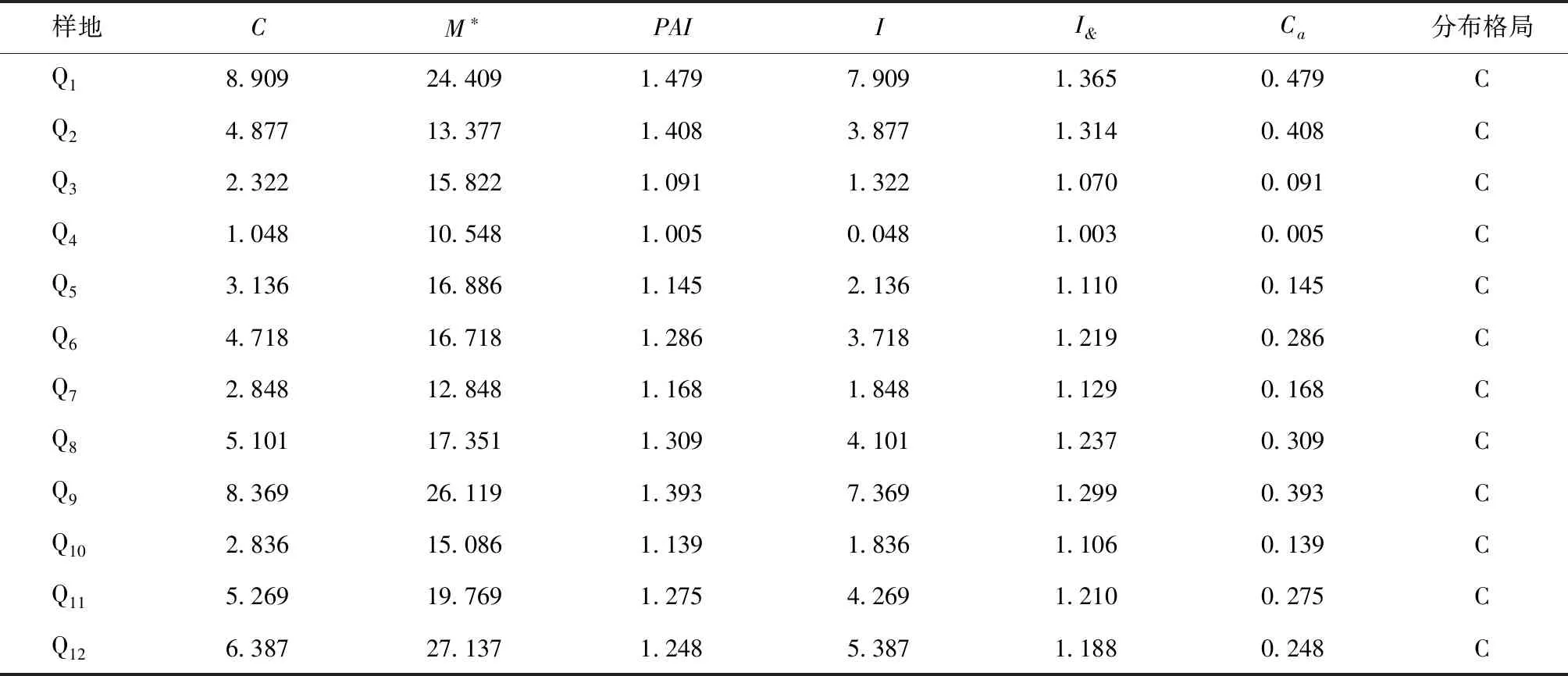
表4 云南松种群空间分布格局Tab.4 The spatial distribution pattern of P.yunnanensis population
3.4 不同龄级空间分布格局
由表5可知,云南松种群Ⅰ—Ⅴ龄级的扩散系数(C)在1.072~5.592之间,均大于1,表明Ⅰ—Ⅴ龄级的云南松种群呈集群分布格局;Ⅵ—Ⅸ龄级的扩散系数(C)在0.636~0.910之间,均小于1,表明Ⅵ—Ⅸ龄级的种群呈均匀分布。其它分布格局指数也得出了同样的结论。由此可见,Ⅵ龄级是云南松种群从集群分布向均匀分布变化的转折点。这表明,随着龄级的增加,云南松种群逐渐由集群分布格局向均匀分布格局转变。
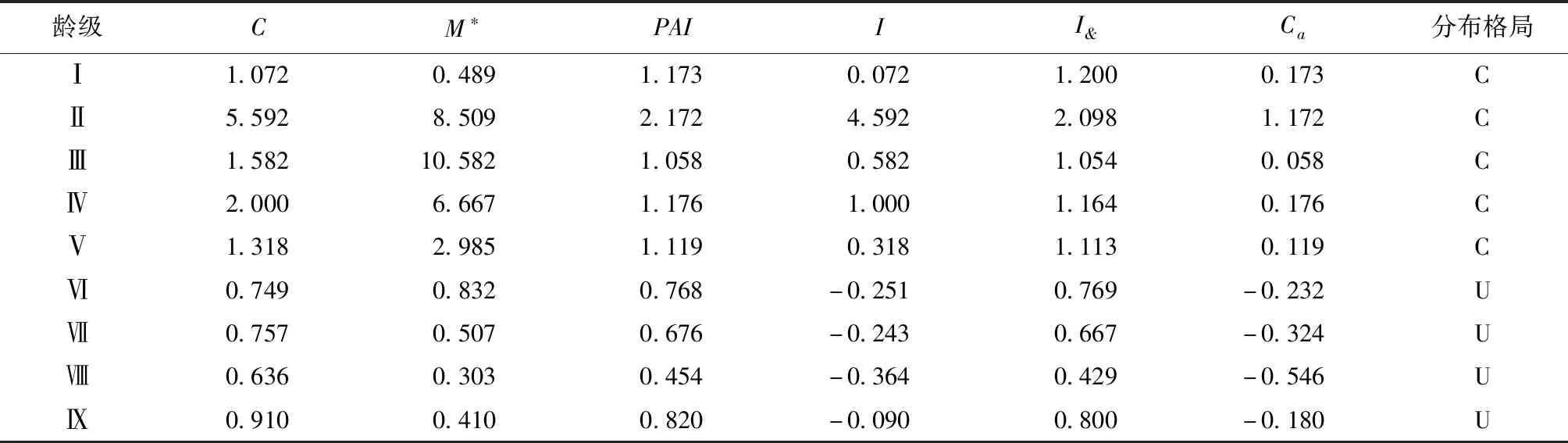
表5 云南松不同径级分布格局Tab.5 Distribution pattern of P.yunnanensis with different diameters
4 讨论
4.1 云南松种群数量动态
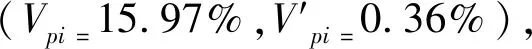
静态生命表分析表明,随着林龄的增加,云南松种群的生命期望值总体上呈下降趋势,在Ⅶ龄级达到最小值;云南松种群死亡率与消失率的变化趋势基本一致,随着龄级的增加,总体上呈上升趋势,在Ⅶ龄级达到最大值。说明云南松种群经过激烈的资源竞争和自疏作用后,Ⅷ龄级个体的生理生态特征更加成熟,抵御外界胁迫的能力和生存能力变强,同时由于种群内个体的大量死亡,释放了一定的生态位,个体间的生存竞争压力减小,生命期望值有所回升,这种“张弛有度”的生存过程有利于云南松种群的繁衍。本研究中云南松种群的存活曲线更趋近于Deevey-II型,但汤明华等[14]对巍山县云南松种群的研究表明其存活曲线为 Deevey-III 型,这可能是由于研究区立地条件的差异造成的。其他学者在不同生物学特性的树种中也发现同一树种的存活曲线在不同研究区也存在一定的差异,例如云杉[21]、辽东栎[27]等,说明种群的存活状况主要受环境条件的影响,其存活曲线并不是固定不变的[29]。
4.2 云南松种群空间分布格局
种群空间分布格局的形成受种群生物学特性、生境异质性、生物间相互作用等多种因素的影响,是不同尺度上各种生物和非生物过程长期相互作用的综合结果[30-31]。已有研究表明,较小尺度上,在物种生物学特性、种子扩散限制和种内与种间竞争排斥等因素的影响下,种群的空间分布常呈聚集分布格局[32]。本研究结果表明,云南松种群空间分布格局为集群分布状态,这与黄小波等[15]的研究结果一致。首先,云南松种子有翅,主要借助于风力进行传播,但由于扩散限制,传播距离不会太远,种子依然散布于母树周围,离母树越远,种子数量越少,导致种群聚集分布;而且由于贵州喀斯特山地生境异质性较高,地面起伏不平,岩石裸露与土壤侵蚀造成的小生境斑块分布的不连续性,对种子的传播产生了阻碍作用,斑块之间的凹处地形土壤较厚易于种子聚集。这与涂洪润等[33]对桂林岩溶石山檵木种群的研究结果一致。其次,大径级个体对周围小生境的改造有利于新个体的存活,小径级个体通过聚集分布能增强对不良外部条件的抵抗能力,维持种群的发展[34];同时云南松种群的聚集分布有利于促进岩溶作用,加快土壤的形成速度,从而有利于其生长繁殖,也导致了云南松个体在小尺度上的聚集分布。所以,扩散限制性、种群自身的生物学特性和所处的生境条件共同作用形成了云南松种群集群分布的空间格局。但是,云南松种群在不同样地间的集聚程度不同,这可能是由于每个种群所处样地的海拔、地形、光照和养分等条件不同所造成的。
4.3 种群发育阶段与空间分布格局
植物为了充分利用有限的资源条件(如光、水、热、营养物质等),种群在不同的发育阶段会表现出不同的空间分布格局[35]。这可能是由种子的散布性[34]、种内与种间的竞争排斥[30]、环境变化和人为干扰等原因造成的。一般认为,随着年龄的增加,种群空间分布格局由集群分布向均匀分布或者随机分布转变[37]。本研究发现,云南松种群在幼龄期和中龄期时(Ⅰ—Ⅴ龄级)呈集群分布,在成年期和老龄期时(Ⅵ—Ⅸ龄级)呈均匀分布。这可能是由于云南松种子受扩散机制影响,散布于母树周围,发育成幼苗,幼苗对资源的需求少且竞争能力薄弱,群体效应有助于缓解外部的竞争压力,促进自身生长,形成了生活史初期的集群分布格局[38]。随着林木的生长发育,植株个体对生存空间和光照、养分、水分等资源的需求增大,受密度制约和有限环境资源的限制,种内、种间竞争加剧出现自疏和它疏现象,导致种群数量个体减少,空间分布格局逐渐趋向于均匀分布或者随机分布[39]。这与刘浩栋等[40]认为随着种群年龄的增加,陆均松种群空间分布格局由集群分布转变为均匀分布的研究结果一致。种群空间分布格局随发育阶段而发生变化,是种群的一种适应机制或者生存策略,有利于植株获取更多的环境资源,提高种群对环境的适应能力[41]。
5 结论与建议
本研究结果表明,云南松种群年龄结构完整,呈纺锤型,主要集中在Ⅲ—Ⅵ龄级;种群数量动态指数分析表明,云南松种群整体表现为增长型,但由于幼苗、幼树不足,种群将难以长期维持稳定。云南松种群空间分布格局为集群分布,不同样地间的集聚程度不同;不同生长阶段云南松种群的空间分布格局不同,随着林龄的增加,云南松种群逐渐由集群分布向均匀分布转变。
因此,在今后云南松林的经营管理中,应从三方面来考虑该地区云南松林的保护与经营,提高森林生态系统功能。第一,通过各种抚育措施(如间伐、砍灌、清理林下活地被物等)改善幼苗的生存环境,减少生长阻力,提高幼苗存活率;第二,开辟林窗,砍掉“老、弱、病、残”个体以及“霸王树”,促进林下幼小个体更多的进入林冠层;第三,加强管理力度,提高民众的科学认识,尽量减少周围村民过度放牧以及踩踏对幼苗生存环境造成的不可逆破坏。
