特有植物厚壳红瘤果茶自然种群生命表及生存力分析
2016-07-23袁丛军杨冰戴晓勇
袁丛军++杨冰++戴晓勇

摘要:为揭示贵州省特有种厚壳红瘤果茶(Camellia rubituberculata)种群结构与分布格局特征,基于模式产地及集中分布区野外踏查资料,选取天然种群集中分布的地段,采用相邻格子法设置典型群落样地,对比分析不同干扰生境下厚壳红瘤果茶自然种群的胸径结构特征,制定静态生命表并对生存力进行分析。结果表明,兴仁县和晴隆县厚壳红瘤果茶萌生、实生种群数量整体上均呈现随着径级结构增大逐渐减小;兴仁县种群以实生种群为主(87.4%),而萌生种群在晴隆县分布地占据重要比重(45.5%);XRM、QLM、XRT种群呈增长型,而XRS、QLS、QLT为衰退型种群。除QLS种群存活曲线呈 Deevey-Ⅲ型外,其余植株种群存活曲线均呈Deevey-Ⅱ型;各种群存活曲线类型方程检验解释量为XRS>QLS>XRM>QLM;XRM、QLS种群死亡率和亏损率均呈现逐渐降低趋势,且降低速率表现为XRM>QLS;QLM、XRS种群死亡率和亏损率均呈现逐渐升高趋势,且升高速率表现为QLM>XRS。种群的生存率总体上逐渐下降,随后保持平稳趋势,不同干扰强度与起源的下降速率及平稳点有所不同。其中,XRM种群到Ⅴ龄级时、QLS种群到达Ⅵ龄级时、晴隆县萌生种群到达Ⅸ龄级时基本达到平稳,而兴仁县实生种群并无明显的平稳点;种群累积死亡率[F(t)]与种群生存率[S(i)]相反。各种群的平均死亡密度率[f(t)]呈现QLM(0.090)>XRS(0.089)>QLS(0.084)>XRM(0.078),而平均危险率[λ(t)]则呈现QLS(0.372)>XRM(0.335)>QLM(0.323)>XRS(0.265)。
关键词:厚壳红瘤果茶;自然种群;静态生命表;存活曲线;贵州省
中图分类号: S685.140.1文献标志码: A文章编号:1002-1302(2016)05-0275-06
厚壳红瘤果茶(Camellia rubituberculata)是山茶科(Theaceae)山茶属(Camellia)常绿小乔木,高度可达8 m,1984年发现于晴隆县,为贵州省特有种[1],主要分布于晴隆县和兴仁县,现有资源量极少,仅分布于贵州省兴仁县田湾乡新寨村和晴隆县紫马乡上捧碧,分布面积共计超过 46.6 hm2。该物种性喜光,叶常绿、花大且花期长(花期10月至翌年3月)、树形优美,可作为优良乡土园林植物优选;木材坚硬、细致,可作为农具、细木工等优良用材优选[2];其叶、油、花在化妆品领域有重要的科研价值[3];因其自然生境极为特殊,该树种适生于喀斯特石山地,可作为石漠化区植被恢复生态经济型乡土树种。然而,有关该种的研究并引起足够的关注,尤其对厚壳红瘤果茶天然种群生命表及生存分析尚未见报道。本研究以典型厚壳红瘤果茶天然群落为研究对象,采用种群生命表及生存分析理论对厚壳红瘤果茶天然种群进行调查研究,揭示该区域厚壳红瘤果茶种群的数量动态变化特征、种群变化趋势,从而为野生厚壳红瘤果茶天然群落的保护、该种资源的示范推广以及在退化生态系统适宜地段上的人工林培育开发利用提供科学的理论依据和指导。
种群不仅是连接群落与个体之间的纽带,也是生物群落、生态系统的基本组成成分。对植物种群研究的最终目的是要阐明种群的动态分化与适应的过程[4]。种群的数量特征是种群的动态分化研究核心内容,包括种群数量、密度、年龄、性比等种群数量统计基本参数,可以反映种群在空间上的变动规律,从而阐释种群动态及其所在群落的演替趋势[5]。种群结构及其变化规律一直是植物生态学的研究热点之一,种群动态研究是种群生态学的核心。植物种群静态生命表和存活曲线的刻画,能够揭示种群生活史中的生死规律和期望寿命,可以为分析珍稀濒危植物种群的保护和珍贵植物速生期利用提供科学依据。通过对种群生命表的编制可得出死亡率、损失率等重要参数,从而为种群数量统计提供更多信息。因此,开展植物种群数量与动态变化规律的研究,对种群的保育管护、预估种群变化趋势和种群资源的开发利用具有重要意义。
1材料与方法
1.1研究区概况
研究区位于贵州省兴仁县田湾乡(新寨村石峩厂组石峩厂)和晴隆县紫马乡(龙头村上捧碧),属于中亚热带季风气候,雨热同期。年均气温为13.9~15.2 ℃,≥10 ℃积温 4 098.0~4 531.2 ℃,极端最低气温为-6.2 ℃,最高气温为21.7 ℃,年日照时间1 513.2 h,无霜期281~290 d,年均降水量1 323.7~1 577.2 mm,4—9月降水量 1 065.1~1 249.7 mm,占全年降水量的80.0%。该区属于喀斯特与砂页岩镶嵌的丘陵地貌,生境为石灰岩山地,岩石裸露率70%~90%。群落为常绿落叶阔叶混交林,其中乔木层主要植物有星毛石栎(Lithocarpus petelotii)、青冈栎(Cyclobalanopsis glauca)、厚壳红瘤果茶(Camellia rubituberculata)、皂荚(Gleditsia sinensis)、云南樟(Cinnamomum glanduliferum)、川黔润楠(Machilus chuanchienensis)、黄连木(Pistacia chinensis)等; 灌木层植物有厚壳红瘤果茶、清香木(Pistacia weinmanifolia)、火棘(Pyracantha fortuneana)、长叶冻绿(Rhamnus crenata)、齿叶铁仔(Myrsine semiserrata)、尖瓣瑞香(Daphne acutiloba);主要藤本植物有大叶酸藤子(Embelia subcoriacea)、杠柳(Periploca sepium)、山橙(Melodinus suaveolens)等;草本植物分布较少。
1.2研究方法
1.2.1野外调查方法在野外实地踏查的基础上,选取特有植物厚壳红瘤果茶种群天然分布区贵州省兴仁县田湾乡、晴隆县紫马乡天然群落为研究对象,分别设置具代表性的样地(30 m×30 m) 3个,共12块样地,总样地面积2 700 m2 。记录每个样地的海拔、坡向、坡度等指标,对样地内物种进行每木调查,调查各物种株高、地径(胸径)、冠幅、树高、枝下高等指标,同时在每个样地中等距设置6个2 m×2 m样方进行灌木层调查、6个1 m×1 m样方进行草本层调查(表1)。
1.2.2龄级划分为避免破坏野生植物资源,本研究采用“空间替代时间”的方法,即将林木依胸径大小分级,以立木级结构代替年龄结构分析种群动态。关于种群大小结构立木级的划分标准,许多学者提出了不同的标准[6-10]。根据该种的生物学特性,本试验以灌丛为单株,采用植株胸径(DBH)作为个体大小的指标研究种群大小结构。径级划分方法:Ⅰ级(DBH<1.0 m)、Ⅱ级(1.0≤DBH<2.0 cm)、Ⅲ级(2.0≤DBH<3.0 cm)、Ⅳ级(3.0≤DBH<4.0 cm)、Ⅴ级(4.0≤DBH<5.0 cm)、Ⅵ级(5.0≤DBH<6.0 cm)、Ⅶ级(6.0≤DBH<7.0 cm)、Ⅷ级(7.0≤DBH<8.0 cm)、Ⅸ级(8.0≤DBH<9.0 cm)。采用陈晓德的数学模型,推导种群年龄结构的动态指数对厚壳红瘤果茶种群动态进行定量描述[11]。
1.2.3生命表编制及生存函数计算由于静态生命表反映多个世代重叠的年龄动态历程中的一个特定时间,而不是对这一种群的全部生活史的追踪,并且调查中存在系统误差,在生命表中会出现死亡率为负的情况,因此本研究采用匀滑技术对数据进行处理。采用生存率函数S(t)、累积死亡率函数F(t)、死亡密度函数f(t)和危险率函数λ(t)等指标对厚壳红瘤果茶自然种群的生存规律及其种群动态进行分析[12]。
2结果与分析
2.1种群起源及年龄结构特征
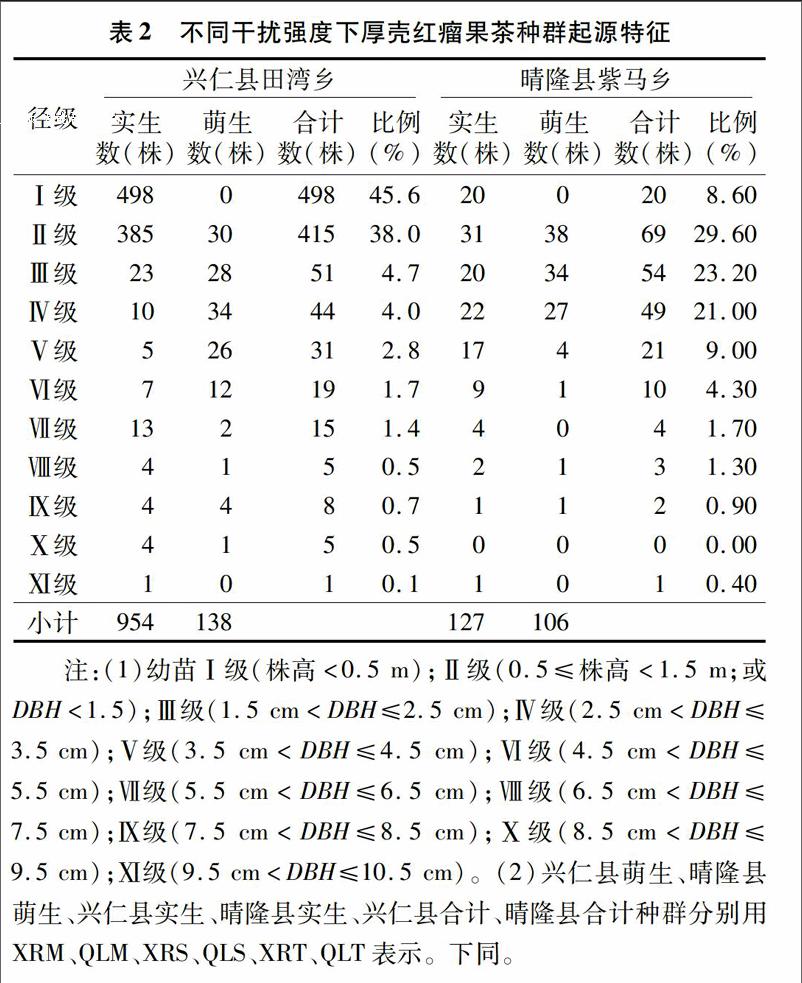
对不同干扰强度下厚壳红瘤果茶种群起源特征统计发现,兴仁县田湾乡厚壳红瘤果茶种群数量显著高于晴隆县紫马乡;其中田湾乡分布的Ⅰ级、Ⅱ级实生幼苗共有883株,萌生幼苗30株,占总植株数的83.6%;而紫马乡分布的Ⅰ级、Ⅱ级实生幼苗共有51株,萌生幼苗38株,占总植株数的 38.2%。不同径级统计表明,实生苗呈现954株(田湾乡)>127株(紫马乡)、萌生苗呈现138株(田湾乡)>106株(紫马乡),而萌生 ∶实生则呈现0.835(紫马乡)>0.145(田湾乡)。研究结果表明,田湾乡(干扰较弱,生境保护水平较好)苗木种子繁育较好,而紫马乡较弱(表2)。
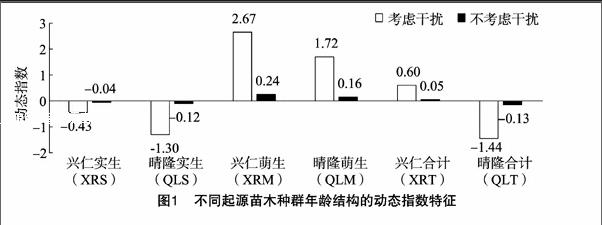
种群的龄级结构是指各龄级组的个体在种群内的比例和配置情况。总体看来,兴仁县和晴隆县厚壳红瘤果茶萌生、实生种群数量均呈现随着径级结构增大逐渐减小。基于量化分析方法绘制不同厚壳红瘤果茶种群龄级结构的动态指数(Vpi)如图1所示,可知:(1)考虑干扰情况下,种群龄级结构的动态指数大小依次为XRM种群(2.67)>QLM种群(1.72)>XRT种群(0.60)>XRS种群(-0.43)>QLS种群(-1.30)>QLT种群(-1.44);(2)不考虑干扰情况,种群龄级结构的动态指数大小依次为XRM种群(0.24)>QLM种群(0.16)>XRT种群(0.05)>XRS种群(-0.04)>QLS种群(-0.12)>QLT种群(-0.13);研究结果表明,XRM、QLM、XRT种群呈增长型,而XRS、QLS、QLT为衰退型种群。
2.2不同种群静态生命表特征
静态生命表是根据一定时期样地内一个种群所有个体的年龄数据编制而成[13],不仅可以反映种群从出生到死亡的数量动态,还可用于预测种群未来发展的趋势。以生存分析理论为基础,编制不同干扰强度下不同起源厚壳红瘤果茶种群静态生命表(表3)。
由表 3可以看出, 不同干扰下实生和萌生种群均呈现种
群数量随着龄级结构的增加逐渐减小,而萌生种群均缺乏Ⅰ龄级种群植株。ex反映了x龄级内个体的生命期望寿命,研究发现,XRS与QLM幼年阶段(Ⅰ~Ⅲ级)表现出高的生命期望,说明此阶段厚壳红瘤果茶植株种群的生存质量较高、生理活动旺盛;随着龄级的增加,植株间的竞争和其他人为干扰影响导致高龄级的生命期望逐渐减小。
2.3不同种群存活曲线特征
存活曲线借助于存活个体数量以描述特定年龄的存活率,反映种群个体在各年龄级的存活状况[14]。本研究以标准化存活量lx为纵坐标、以径级相对的龄级作横坐标,绘制了存活曲线(表4)。由此可知,除晴隆县实生植株种群存活曲线呈Deevey-Ⅲ型外,其余植株种群存活曲线均呈Deevey-Ⅱ型;各不同起源厚壳红瘤果茶植株种群存活曲线类型方程检验解释量依次为兴仁县实生(R2=0.963)>晴隆县实生(R2=0.935)>兴仁县萌生(R2=0.829)>晴隆县萌生(R2=0.815)。
2.4死亡率和亏损率曲线
以厚壳红瘤果茶不同种群的龄级为横坐标,死亡率(qx)和亏损率(Kx)为纵坐标,绘制该种群的死亡率和亏损率曲线(图2)。由图 2可知,厚壳红瘤果茶各种群的死亡率和亏损率曲线的变化基本一致。总体看来,XRM、QLS种群死亡率和亏损率均呈现逐渐降低趋势,且降低速率表现为XRM>QLS;QLM、XRS种群死亡率和亏损率大致呈现逐渐升高趋势,且升高速率表现为QLM>XRS。
2.5种群生存函数分析
厚壳红瘤果茶植株种群的生存率总体上逐渐下降,随后保持平稳趋势,不同干扰强度与起源的下降速率及平稳点有所不同(图3)。其中,兴仁县萌生种群到Ⅴ龄级时、晴隆县实生种群到达Ⅵ龄级时、晴隆县萌生种群到达Ⅸ龄级时基本达到平稳,而兴仁县实生种群并无明显的平稳点;与之相应,种群累积死亡率与种群生存率相反。各种群的平均死亡密度呈现QLM(0.090)>XRS(0.089)>QLS(0.084)>XRM(0078),而平均危险率则呈现QLS(0.372)>XRM(0.335)>QLM(0.323)>XRS(0.265)。XRM种群死亡密度表现为先保持不变(Ⅰ~Ⅲ级),后迅速减小并保持不变(Ⅴ~Ⅸ级),随后在Ⅹ级处出现小波峰;QLM种群死亡密度则表现为先在Ⅱ级处出现小波峰,后保持不变(Ⅲ~Ⅷ级),后逐渐迅速减小并在Ⅹ级处又出现小波峰;XRS种群死亡密度表现为先保持不变(Ⅰ~Ⅲ级),随后即在Ⅳ处出现小波峰,后保持不变(Ⅳ~Ⅸ级),随后在Ⅹ级处出现小波峰;QLS种群死亡密度则表现为先保持不变(Ⅰ~Ⅲ级),后先减小后迅速增
加,并在Ⅴ级处出现小波峰,后保持不变(Ⅵ~Ⅸ级),随后在Ⅹ级处出现小波峰。而XRM种群危险率表现为先增加后减小,再保持平稳不变(Ⅵ~Ⅹ级);QLM种群危险率表现为先增加后减小;XRS种群危险率表现为先增加后减小,随后又
增加趋势;QLS种群则表现为先增加后减小,后又呈现先增加后减小,最后又再次呈现增加趋势。
3结论与讨论
3.1结论
兴仁县和晴隆县厚壳红瘤果茶萌生、实生种群数量均随着径级结构增大逐渐减小;其中,兴仁县种群以实生种群(种子繁育)为主(87.4%),而萌生种群在晴隆县分布地占据重要比重(45.5%);XRM、QLM、XRT种群呈增长型,而XRS、QLS、QLT为衰退型种群。
除QLS种群存活曲线呈Deevey-Ⅲ型外,其余植株种群存活曲线均呈Deevey-Ⅱ型;各种群存活曲线类型方程检验解释量为XRS>QLS>XRM>QLM;XRM、QLS种群死亡率和亏损率均呈现逐渐降低趋势,且降低速率表现为XRM>QLS;QLM、XRS种群死亡率和亏损率均呈现逐渐升高趋势,且升高速率表现为QLM>XRS。
厚壳红瘤果茶植株种群的生存率总体上逐渐下降,随后保持平稳趋势,不同干扰强度与起源的下降速率及平稳点有所不同。其中,兴仁县萌生种群到Ⅴ龄级时、晴隆县实生种群到达Ⅵ龄级时、晴隆县萌生种群到达Ⅸ龄级时基本达到平稳,而兴仁县实生种群并无明显的平稳点;与之相应,种群累积死亡率与种群生存率相反。
3.2讨论
植物地理学研究中,常常将分布狭限或是异常狭限分布区的物种或其他分类单元称为特有植物[1]。贵州省特有植物是仅分布在贵州省地理区域内,而其他地区没有的植物种类(目前未见报道或发现),其起源、发育演化和传播呈间断或不间断狭限分布在贵州省地理区域内。山茶属植物是贵州省特有植物种类最多的属,约分布有16或21种[15-16],作为贵州省特有植物(280种)的重要组成部分,其表征着贵州省特殊生境条件下植物适应进化演变的结果。贵州省特有植物的保护与利用作为区域生物多样化保护过程中一项极其重要研究内容和热点,厚壳红瘤果茶隶属山茶科山茶属瘤果茶组常绿小乔木,是典型的生长于岩溶区山旮旯地段乡土适生油脂兼园林观赏植物;分布区狭窄,资源量较少,多散生于常绿落叶阔叶林中,群落建群种以壳斗科植物为主,该种为伴生种[17-18]。因此,开展以野生自然种群动态变化和现有种群生存力分析具有重要意义。
种子繁育(实生苗)和萌蘖繁育(萌生苗)作为种子植物种群天然更新的2种重要方式,种群采取何种生存对策取决于该物种的生物学特性和外界环境的压力影响[19-20]。前人对特有植物种群动态变化开展了许多研究工作[19,21-24],而从种群不同起源方式及干扰型对种群自然更新研究工作较少[25-26]。本研究对瘤果茶组(Sect. tuberculate)贵州省特有典型生态经济型厚壳红瘤果茶实生与萌生植株种群特征进行研究表明,该种种群繁育以实生苗为主(XRS、QLS所占比例分别为87.4%、54.5%),QLM种群数量是QLT种群数量的重要组成部分(45.5%)。晴隆县紫马乡分布点由于种群资源量少,资源分布面积较小,且群落及生境地受人为干扰破坏较严重,萌生植物种群所占比例较大,成为该地厚壳红瘤果茶种群更新繁育的重要组成部分;兴仁县田湾乡分布点由于种群资源量较大,资源分布面积较大,且群落及生境地受人为干扰破坏较弱,实生植株种群占据了大量的比重(87.4%),人为干扰对种群更新,尤其是特有植物的种群更新具有显著影响。研究结果与袁丛军等对贵州省特有植物岩生红豆研究结果一致,强干扰型显著影响了目的树种的种群更新与繁育[27]。然而有关强干扰型如何影响厚壳红瘤果茶种群更新生态学过程的研究有待进一步深入和开展。
植物种群的生命表绘制和编制,反映该种群生存现状,对揭示植物种群与环境适应的结果和阐明种群或群落未来发展趋势具有重要意义[28]。由于现有科研试验条件限制,要完全追踪种群从发生在最后一个个体死亡的真实过程,获取植物种群动态生命表是一项极其困难的工作,尤其是针对一些乔木长寿命植物种群而言[29]。生存分析理论用于研究种群的动态和结构变化,在生命表分析中具有较高的实际应用价值,能更合理、直观地说明种群的结构和动态变化[30-31]。本研究鉴于该种群的自身生物学特性和生境特征,结合前人有关山茶属植物研究结果[32-33],研究采用以1 cm为1个径级单位对不同种群划分龄级,编制不同起源和干扰情况下的植物种群生命表[34],并对各种群进行生存力分析。研究结果表明,QLS种群存活曲线呈Deevey-Ⅲ型(凹曲线型),说明晴隆县实生种群在幼苗早期死亡率高,但一旦活至某一年龄,种群个体死亡率就会处于较低水平;而QLM、XRS、XRM种群存活曲线均呈Deevey-Ⅱ型(直线型),该类种群各年龄的死亡率基本一致。结合研究区的人为干扰活动情况,研究认为强干扰型的人为活动极大促进厚壳红瘤果茶实生植株种群的幼龄(Ⅱ级→Ⅲ级)苗木的死亡数量及速率,对萌生植株种群(QLM种群死亡数量和速率)影响不显著;弱干扰型的人为活动对XRM植株种群(Ⅰ级→Ⅵ级)影响显著强于XRS种群。因此,建立保护小区并减少人为活动强度对厚壳红瘤果茶野生种群的干扰,可有效保护野生实生种群,丰富和保持更多的遗传多样性。
参考文献:
[1]林蒙嘉,卢其明. 贵州山茶新纪录[J]. 中山大学学报:自然科学版,1984(2):83-85.
[2]邓忠治,潘德权,刘正本,等. 贵州省厚壳红瘤果茶初步研究[J]. 贵州林业科技,2011(2):39-42.
[3]周丽,王苑,张应祝,等. 厚壳红瘤果茶果实特征与种子萌发特性[J]. 北方园艺,2015(2):34-37.
[4]姜汉侨,段昌群,杨树华,等. 植物生态学[M]. 北京:高等教育出版社,2010.
[5]张金屯,吴冬丽,张斌,等. 北京地区几种重点保护植物生存群落生态学研究[M]. 北京:中国科学技术出版社,2013.
[6]张治国,王仁卿. 中国分布北界的山茶 (Camellia japonica)种群大小结构和空间格局分析[J]. 植物生态学报,2000(1).
[7]洪伟,王新功,吴承祯,等. 濒危植物南方红豆杉种群生命表及谱分析[J]. 应用生态学报,2004(6):1109-1112.
[8]茹文明,张桂萍,毕润成,等. 濒危植物脱皮榆种群结构与分布格局研究[J]. 应用与环境生物学报,2007(1):14-17.
[9]闫淑君,洪伟,吴承祯,等. 丝栗栲种群生命过程及谱分析[J]. 应用与环境生物学报,2002(4):351-355.
[10]吴承祯,洪伟,谢金寿,等. 珍稀濒危植物长苞铁杉种群生命表分析[J]. 应用生态学报,2000(3):333-336.
[11]陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报,1998(2):104-107.
[12]卢杰,潘刚,罗大庆,等. 濒危植物急尖长苞冷杉种群生命表分析[J]. 水土保持研究,2010(5):212-216.
[13]吴承祯,洪伟,谢金寿,等. 珍稀濒危植物长苞铁杉种群生命表分析[J]. 应用生态学报,2000(3):333-336.
[14]李玲,张光富,王锐,等. 天目山自然保护区银杏天然种群生命表[J]. 生态学杂志,2011(1):53-58.
[15]屠玉麟. 贵州特有植物初步研究(一)[J]. 贵州林业科技,1991(3):68-81.
[16]张华海,班平原. 贵州山茶属植物地理分布及开发利用[J]. 贵州科学,2007(1):68-72.
[17]邹天才,刘海燕,周洪英,等. 贵州重要野生观赏植物的引种繁殖和应用栽培技术研究[J]. 种子,2007,26(6):12-15,20.
[18]邓忠治,潘德权,刘正本,等. 贵州省厚壳红瘤果茶初步研究[J]. 贵州林业科技,2011(2):39-42.
[19]刚群,闫巧玲,刘焕彬,等. 种子更新与萌蘖更新蒙古栎一年生幼苗生长特性的比较[J]. 生态学杂志,2014(5):1183-1189.
[20]李小双,彭明春,党承林. 植物自然更新研究进展[J]. 生态学杂志,2007(12):2081-2088.
[21]龚滨,夏洋洁,张光富,等. 中国特有珍稀濒危树种银缕梅种群结构和空间格局[J]. 生态与农村环境学报,2012,28(6):638-646.
[22]秦凤,邓洪平,由永飞,等. 重庆市特有植物缙云卫矛的种群动态[J]. 贵州农业科学,2011(4):27-31.
[23]袁春明,孟广涛,方向京,等. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布[J]. 生态学报,2012(12):3866-3872.
[24]杨乃坤,邹天才,刘海燕,等. 贵州特有植物长柱红山茶种群年龄结构及空间分布格局研究[J]. 热带亚热带植物学报,2015(2):205-210.
[25]王赛宵,李清河,徐军,等. 克隆植物唐古特白刺不同层次的种群结构与生命表分析[J]. 东北林业大学学报,2013,41(1):72-76.
[26]郭有燕,周建云,何景峰,等. 文冠果种群繁殖方式及其在种群更新中的作用[J]. 应用生态学报,2014(11):3110-3116.
[27]袁丛军,安明态,严令斌,等. 两种干扰强度下岩生红豆天然群落特征及物种多样性分析[J]. 南方农业学报,2013(2):280-284.
[28]张悦,易雪梅,王远遐,等. 采伐对红松种群结构与动态的影响[J]. 生态学报,2015(1):38-45.
[29]何亚平,费世民,蒋俊明,等. 不同龄级划分方法对种群存活分析的影响——以水灾迹地油松和华山松种群生存分析为例[J]. 植物生态学报,2008(2):448-455.
[30]洪伟,王新功,吴承祯,等. 濒危植物南方红豆杉种群生命表及谱分析[J]. 应用生态学报,2004,15(6):1109-1112.
[31]李建贵,潘存德,彭世揆,等. 天山云杉种群统计与生存分析[J]. 北京林业大学学报,2001(1):84-86.
[32]詹明勋,何坤益,徐震骐,等. 台湾中部暖温带阔叶林年轮特征、生长与树龄结构[J]. 北京林业大学学报,2015(3):84-93.
[33]杜兴乔,刘映良. 荔波瘤果茶种群生命表分析[J]. 安徽农业科学,2009(33):16613-16615.
[34]谢佳燕,林佳. 亚致死浓度吡虫啉对禾谷缢管蚜实验种群生命表参数的影响[J]. 江苏农业科学,2014,42(5):125-126.何遵卫,周章义,马凤翔. 4个频率下不同生长等级国槐的电生理分析[J]. 江苏农业科学,2016,44(5):281-283.
