反复冻融过程中开背调味鱼品质 及蛋白特性的变化
2021-06-03吴晨燕王珂莉刘鑫洁刘思佳马俪珍
吴晨燕,杨 梅,王珂莉,刘鑫洁,刘思佳,马俪珍,*
(1.天津农学院食品科学与生物工程学院,国家大宗淡水鱼加工技术研发分中心(天津),天津 300384;2.天津市强源食品有限公司,天津 301700)
水产品保鲜包括冰藏保鲜、冷海水保鲜、微冻保鲜[1]、 冻结保鲜[2]、超冷保鲜、气调保鲜、化学保鲜等方式。其中,冷冻保鲜不仅能延长产品保质期,且经济成本较低,是一种比较常见的保鲜方式。冷冻保鲜通过将鱼体的水分冻结成冰,使其体内的生化反应和微生物活动基本处于停滞状态[3-4],从而达到延长鱼肉货架期的目的[5-6]。 随着冷链物流业的发展,冷冻品在-18 ℃环境下由生产地配送至全国各地也成为可能,但根据实际生产情况,冷冻水产品从厂家库房装载至物流车、物流车运输、从物流车卸货至目的地仓库房、消费者购买产品带回家这一系列过程中都伴随着因温度波动而导致产品品质下降的情况。鱼肉在经历冷冻-解冻循环过程中,其蛋白质发生不同程度的变性。研究发现,随着冷冻-解冻循环次数的增加,斑节对虾和南美白对虾[7]、尖吻鲈 鱼肉[8]、脆肉鲩鱼肉[9]等都存在感官品质和风味下降、蛋白和脂肪氧化、组织结构和功能特性变化等现象。
随着市场上调理食品的兴起,调味鱼产品也越来越受到消费者的喜爱,因调味鱼产品的保鲜方式以冷冻保鲜更为常见,调味后的鱼肉产品在配送转运过程中也伴随着反复冻融过程,关于此过程产品品质变化的研究尚不深入。本实验以调味开背斑点叉尾鮰为研究对象,将其速冻后在-18 ℃下冻藏,通过超高压(ultra high pressure,UHP)和滚揉(tumbling,TB)两种腌制方式制得开背调味鱼产品,测定调味鱼肉在5 次冻融循环过程中菌落总数、硫代巴比妥酸反应物(thiobarbituric acid reaction substances,TBARS)、总挥发性盐基氮(total volatile basic nitrogen,TVB-N)含量及肌原纤维蛋白(myofibril protein,MP)相关指标,并通过扫描电子显微镜、紫外分光光度计、傅里叶变换红外光谱(Fourier transform infrared spectroscopy,FTIR)仪对MP的结构进行表征,以探究在冻藏过程中反复冻融循环次数对调味鱼品质、结构和功能特性的影响,为开背调味鱼新产品投入市场提供一定的数据支持和参考。
1 材料与方法
1.1 材料与试剂
斑点叉尾鮰、香辛料、调味料、姜、葱、蒜购于天津市西青区红旗农贸水产批发市场。
一水柠檬酸(食品级) 常州百运渡化工有限 公司;五香浓缩液 顶兴(天津)食品科技发展有限 公司;硝酸银、盐酸 国药集团化学试剂有限公司。以上试剂均为分析纯。
1.2 仪器与设备
HPP.L1-600/5超高压设备 天津市华泰森淼生物工程技术有限公司;BVRJ-40真空滚揉机 嘉兴艾博实业有限公司;FA25高速剪切分散乳化机 德国弗鲁克 公司;Pro台式扫描电子显微镜 荷兰Phenom-Word BV公司;MinI 4电泳仪 美国Bio-Rad公司;UV-1800紫外分光光度计 日本岛津公司;Spectrum65 FTIR仪 美国PerkinElmer公司。
1.3 方法
1.3.1 斑点叉尾鮰鱼处理
购买鲜活斑点叉尾鮰鱼,经宰杀(分左右两半胴体)、减菌化处理后,采用超高压(UHP组)和滚揉(TB组)方式腌制鱼体,将腌制好的鱼体真空包装后放入速冻机(-35 ℃)至中心温度达-18 ℃(大约30 min),然后在-18 ℃冰箱冷冻贮藏,冻藏12 h后测定新鲜鱼肉指标,此时冻融循环次数记为0(不同处理样品分别记为UHP0d和TB0d)。总冻藏时间为100 d,每组样品冻结数量为18 袋,期间每隔20 d,将全部样品取出,置于流动水中解冻,直至鱼体中心温度达0~4 ℃,即完成第一次冻融循环(不同处理样品分别记为UHP20d和TB20d),此时每组样品留下3 袋用于指标的测定,其余样品立即放入-18 ℃冰箱中继续冻藏,20 d后再次将冰箱中的所有样品从冰箱中取出,进行相同方式的冻融循环,以此类推进行第2、3、4、5次的冻融循环(UHP组中分别记为UHP40d、UHP60d、UHP80d和UHP100d,TB组中 分别记为TB20d、TB40d、TB60d、TB80d、TB100d)。每次冻融循环的样品进行各项指标的测定。
超高压腌制条件:食盐质量分数11%、超高压压力200 MPa、保压时间15 min。
滚揉腌制条件:食盐质量分数10%、滚揉方式为呼吸式、真空度0.08 MPa、滚揉时间60 min。此处呼吸式滚揉的具体步骤是:抽真空正转5 min→静置2.5 min→反转5 min→静置2.5 min→正压正转5 min→静置2.5 min→正压反转5 min→静置2.5 min,再一次重复以上过程。
1.3.2 菌落总数的测定
菌落总数测定参考GB 4789.2—2016《食品安全国家标准 食品卫生微生物学检验 菌落总数测定》[10]。
1.3.3 TBARS值的测定
TBARS值测定参考马俪珍等[11]的方法。
1.3.4 TVB-N含量的测定
准确称取5 g绞碎的肉样,置于锥形瓶中,加入50 mL蒸馏水,搅拌均匀,瓶口包上保鲜膜放入摇床振荡30 min,过滤后滤液放入冷藏柜中备用。待半微量定氮蒸馏装置加热沸腾后,吸取5 mL滤液(以蒸馏水作为空白)加入凯氏蒸馏装置反应室,再立即加入5 mL 10 g/L MgO溶液,塞住塞子,加蒸馏水液封。在反应装置末端处用体积分数2%的硼酸作为接收液,液面以下接收5 min,液面以上接收1 min,之后用0.01 mol/L盐酸标准溶液进行滴定,按式(1)计算TVB-N含量。

式中:V1表示样液消耗盐酸体积/mL;V2表示空白消耗盐酸体积/mL;cB表示标准盐酸浓度/(mol/L);m表示样品质量/g;V4表示样液体积/mL;V3表示样液定容后的体积/mL;14表示氮的摩尔质量/(g/mol)。
1.3.5 电子鼻分析气味成分
称取10 g剪碎鱼肉(精确到0.001 g),放入样品瓶中,待上机检测。先用空气清洗传感器,清洗时间为300 s,然后通过真空泵将样品中的气体吸入到电子鼻中,进气速率为0.8 L/min,检测时间为200 s。每种鱼肉样品5 个平行,每个样品测定一次。
1.3.6 MP相关指标的测定
1.3.6.1 MP的提取
参照Xiong Youling L.等[12]的方法进行鮰鱼MP的提取,提取过程在4 ℃条件下操作。制备好的MP采用双缩脲法测定MP的蛋白质量浓度,并放置于4 ℃的冰箱内备用。
1.3.6.2 总巯基含量的测定
总巯基含量的测定参考李鹏[13]的方法。在10 mL离心管中加入0.5 mL 2 mg/mL MP溶液、2.5 mL含8 mol/L 尿素的Tris-甘氨酸溶液(pH 8,每升Tris-甘氨酸溶液中含有10.4 g Tris、6.9 g甘氨酸、1.2 g 乙二胺四乙酸 (ethylene diamine tetraacetic acid,EDTA))以及0.02 mL Ellman’s试剂,再加入20 μL 5,5’-二硫代双 (2-硝基苯甲酸)(5,5’-dithiobis-(2-nitrobenzoic acid),DTNB)避光室温反应25 min,立即在412 nm波长处测定吸光度(A412nm)。利用公式(2)计算总巯基含量。
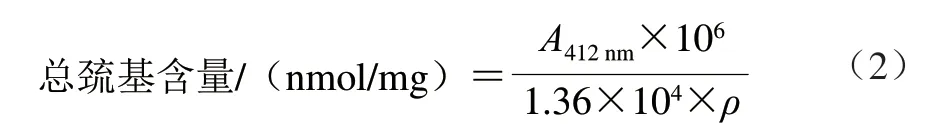
式中:1.36×104表示摩尔吸光系数/(L/(mol·cm));ρ为双缩脲法测得的蛋白质量浓度/(mg/mL)。
1.3.6.3 二硫键含量的测定
二硫键含量的测定参照Thannhauser等[14]的方法。先取1 mol/L Na2SO310 mL,与100 mg DTNB充分混合,调节pH值至7.5,在38 ℃条件下通入氧气直至溶液颜色从亮红色变为淡黄色,大约45 min,得到2-硝基-5-硫 代苯磺酸酯(2-nitro-5-thio- sulfobenzoate,NTSB)储备液;NTSB使用液现配现用。称取3.755 g甘氨酸、0.88 g EDTA、12.6 g Na2SO3、236.32 g硫氰酸胍,用蒸馏水溶解并定容至1000 mL,再将NTSB储备液与混合溶液按体积比1∶100稀释,调节溶液pH值至9.5,即得NTSB使用液。在0.25 mL MP溶液中加入3 mL新制备的NTSB使用液漩涡振荡均匀。将混合溶液避光室温下放置25 min后测定其在412 nm波长处的吸光度,用摩尔吸光系数13900 L/(mol·cm)计算二硫键含量。
1.3.6.4 紫外光谱测定
以0.6 mol/L KCl缓冲液为空白,在230~350 nm波长范围内建立基线,将MP用0.6 mol/L KCl溶液稀释至蛋白质量浓度为1 mg/mL,采用UV-1800紫外分光光度计在230~350 nm波长范围内对MP溶液进行紫外光谱扫描。使用Origin 8.5软件计算二阶导数紫外光谱[15]。
1.3.6.5 FTIR测定
参考喻亚丽[16]的方法,将提取的MP冻干,取冻干样品1 mg,经KBr压片,用Spectrum65 FTIR仪对样品进行红外扫描。扫描范围为1700~1000 cm-1。
1.3.6.6 扫描电子显微镜观察微观结构
将提取的MP真空冷冻干燥成冻干粉,取少量冻干粉均匀涂抹在胶垫上,镀金处理60 s,用扫描电子显微镜观察其微观结构。测定参数:加速电压为10 kV,束流强度为能谱点扫,探头模式为背散射,实时观察参数为684×684 像素,照片存储参数为1024×2048 像素,放大1000 倍。
1.3.6.7 SDS-PAGE分析
参照Laemmli等[17]的方法,在0.25 mL 2 mg/mL MP溶液中分别加入0.25 mL、体积分数10%β-巯基乙醇(β-mercaptoethanol,β-ME)和0.25 mL十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)样品缓冲液(含4 g/100 mL SDS、20 mL/100 mL甘油、0.125 mol/L Tris-HCl,pH 6.8),使MP质量浓度为1 mg/mL,沸水浴中加热3 min后置于冰中冷却,加入50 μL溴酚蓝,以10000×g离心3 min。吸取上清液20 μL进行点样,浓缩胶质量分数为4%,分离胶质量分数为12%。样品在浓缩胶中电流为20 mA,进入分离胶后,电流改为40 mA。
1.4 数据统计与分析
每组实验重复3 次,利用Excel软件统计数据,结果以平均值±标准差表示,采用Simplot 10.0软件作图,用SPSS 13.0软件中的Tukey HSD进行差异显著性分析 (P<0.05表示差异显著)。
2 结果与分析
2.1 反复冻融过程中UHP和TB样品菌落总数的变化

图 1 反复冻融过程中UHP和TB样品菌落总数的变化Fig. 1 Changes in TBC of UHP and TB samples during repeated freeze-thaw cycles
从图1可以看出,UHP和TB样品初始菌落数分别为3.41(lg(CFU/g))和4.86(lg(CFU/g)),前者显著低于后者,而且在冻藏100 d、反复冻融5 次的过程中始终为UHP样品菌落总数显著低于TB样品菌落总数。这是因为超高压处理不仅能够提高腌制速率,还具有冷杀菌作用[18],尤其对芽孢菌有很显著的杀灭作用[19]。UHP和TB样品在冻融1 次后细菌总数分别为3.11(lg(CFU/g)) 和4.38(lg(CFU/g)),相比初始菌落总数均有所下降,这是因为加工好的产品经过速冻、-18 ℃冻藏后,微生物进入新的冷藏环境,在适应及滞留期部分被冻死,这与李秀霞等[20]的研究结果一致。冻融1 次后,随着冻融循环次数的继续增加,UHP和TB样品的菌落总数一直维持在较低水平,并没有显著增加,这是因为在冻藏条件下,微生物细胞内原生质或胶体脱水,溶质浓度增加,促使菌体蛋白质变性,同时冰晶体的形成还会使微生物细胞膜遭受机械性破坏。
2.2 反复冻融过程中UHP和TB样品TBARS值的变化
从图2可以看出,冻融初期,UHP和TB两组样品的TBARS值分别为0.04 mg/kg和0.06 mg/kg,两者均处于 较低水平,随着冻融循环次数增加,TBARS值在冻融前3 次整体上变化不显著(P>0.05),3 次后有显著升高趋势 (P<0.05),冻融4 次时,UHP、TB两组样品的TBARS值分别为0.065 mg/kg和0.074 mg/kg,前者显著低于后者 (P<0.05),这是因为超高压处理具有抑制肉类脂肪氧化的作用[21]。即使冻融5 次,UHP、TB两组样品的TBARS值也仅分别升高至0.10 mg/kg和0.11 mg/kg,且两组之间差异不显著(P>0.05)。这说明UHP和TB产品在冻藏过程中脂肪氧化得到很好的控制,这可能与五香风味腌制液中姜、蒜、香辛料等所含有的天然抗氧化成分有关。

图 2 反复冻融过程中UHP和TB样品TBARS值的变化Fig. 2 Changes in TBARS values of UHP and TB samples during repeated freeze-thaw cycles
2.3 反复冻融过程中UHP和TB样品TVB-N含量的变化
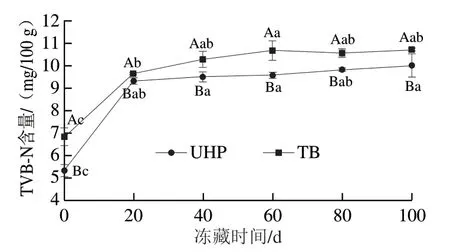
图 3 反复冻融过程中UHP和TB样品TVB-N含量的变化Fig. 3 Changes in TVB-N contents of UHP and TB samples during repeated freeze-thaw cycles
TVB-N含量是评价水产制品新鲜程度的重要指标之一。TVB-N含量与微生物数量高度相关,微生物作用下水产品中的蛋白质被分解产生氨及胺类等碱性含氮 物质[22],使TVB-N含量升高,产品新鲜度下降。从图3可以看出,冻融初期,UHP、TB两组样品TVB-N含量分别为5.34 mg/100 g和6.85 mg/100 g,含量较低,说明这两种腌制方式加工的开背调味鱼新鲜度比较高,这与菌落总数结果一致。冻融循环1 次后,两组TVB-N含量迅速升高,随着反复冻融循环次数的继续增加,TVB-N含量呈缓慢升高趋势,到冻融循环5 次,UHP、TB样品的TVB-N含量分别为10.02、10.73 mg/100 g,均低于淡水鱼鲜度TVB-N含量的限量(20.00 mg/100 g)[23]。这是因为冻藏过程中微生物作用受到抑制,蛋白质被分解为碱性含氮物质的过程也被抑制,即使后期冻融循环次数增加, 因微生物作用导致的TVB-N含量增加量也较小。在冻融循环过程中,UHP样品的TVB-N含量始终显著低于TB样品(P<0.05)。综上,对于冻融5 次的开背调味鱼,超高压腌制比滚揉腌制能更好地保持其鲜度。
2.4 反复冻融过程中UHP和TB样品气味的变化
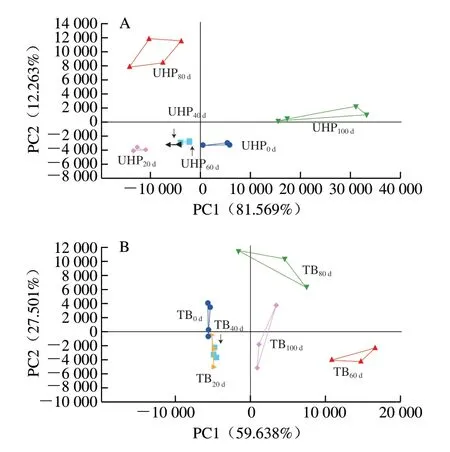
图 4 反复冻融过程中UHP(A)和TB(B)样品气味的主成分分析Fig. 4 Principal component analysis plots for odor changes of UHP (A) and TB (B) samples during repeated freeze-thaw cycles
从图4A可知,冻融循环0~3 次的UHP0d、UHP20d、UHP40d、UHP60d样品较密集地分布在一起,而UHP80d、UHP100d样品与UHP0d样品分布距离较远,说明当将样品冻融循环4 次后,UHP样品的风味与新鲜开背调味鱼之间出现显著差异。从图4B可以看出,TB样品中,TB60d、TB80d、TB100d样品分布在不同的区域,与TB0d样品相距较远,说明当将样品冻融循环3 次后,TB样品的风味与新鲜开背调味鱼之间出现显著差异。以上结果与TBARS值结果基本一致,说明脂肪氧化对调理鱼产品风味影响较大。
2.5 反复冻融过程中UHP、TB 样品MP总巯基和二硫键含量的变化
巯基是蛋白分子中非常重要的功能基团,总巯基含量是蛋白氧化的一项重要指标,总巯基含量越低,蛋白氧化程度越高[24]。总巯基含量的降低是由于巯基氧化为二硫键[25],所以总巯基含量降低对应着二硫键升高。
由图5可以看出,随着冻融循环次数增加,UHP和TB鱼肉产品蛋白氧化程度逐渐加强,表现为MP总巯基含量显著降低(P<0.05),二硫键含量显著升高 (P<0.05)。这是因为反复冻融过程中,冰晶体逐渐增大破坏了细胞结构,蛋白质周边分散的粒子浓度继续增加,加速蛋白质变性,蛋白质发生变性和聚集,会伴随着巯基氧化形成二硫键[26-28]。在冻融循环0 次时,UHP、TB样品的MP总巯基含量较高,分别为93.10、72.17 nmol/mg,二硫键含量较低,分别为26.88、29.11 nmol/mg。但冻融循环5 次后,总巯基含量显著降低,二硫键含量显著升高,其中UHP、TB总巯基含量分别为56.88、51.55 nmol/mg,二硫键含量分别为35.42、44.00 nmol/mg。UHP和TB样品在各个冻融循环次数之间的MP总巯基含量和二硫键含量差异均达到显著水平 (P<0.05),表明超高压腌制加工的开背调味鱼蛋白氧化程度显著低于滚揉腌制方式,这是因为滚揉腌制方式会伴随着产热产能的过程,物理状态也有会发生一定程度的改变,而超高压方式对产品结构影响较小,所以在反复冻融过程中,超高压腌制调味鱼产品的蛋白氧化程度低于滚揉腌制方式。
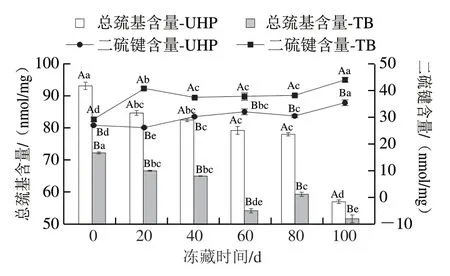
图 5 反复冻融过程中UHP、TB样品MP总巯基和二硫键含量的变化Fig. 5 Changes in total sulfhydryl group and disulfide bond contents of UHP and TB samples during repeated freeze-thaw cycles
2.6 反复冻融过程中UHP、TB样品MP的二阶导数紫外光谱变化
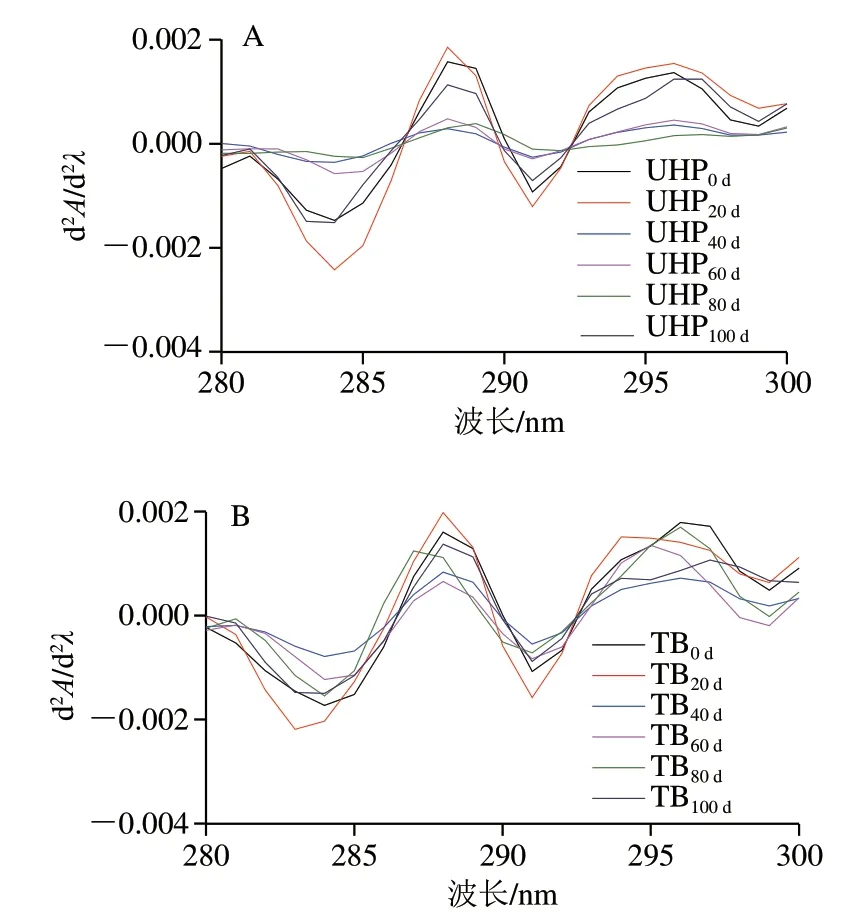
图 6 反复冻融过程中UHP(A)、TB(B)样品MP的 二阶导数紫外光谱变化Fig. 6 Change in second-derivative UV spectra of UHP (A) and TB (B) samples during repeated freeze-thaw cycles
蛋白质分子中的部分官能团具有特征性的紫外吸收区域[29],色氨酸、酪氨酸的R基团含有苯环共轭双键,分别在280 nm和275 nm波长处有吸收峰[30]。从图6可以看出,在280~300 nm之间,不同冻融循环次数UHP和TB样品MP的二阶导数吸收光谱均在288、296 nm波长处出现极大值,在284、292 nm波长处出现极小值。在288 nm波长处的峰值是色氨酸、酪氨酸共同作用的结果,而在296 nm波长处的峰值可以单独归因于酪氨酸[31]。随着冻融循环次数的增加,288 nm和296 nm处的峰值有明显的蓝移,这说明色氨酸、酪氨酸残基从内部转移到极性更高的区域,并由于构象的改变而暴露在蛋白的表面,这表明蛋白发生了一定程度的氧化。
2.7 反复冻融过程中UHP和TB样品MP的FTIR分析结果
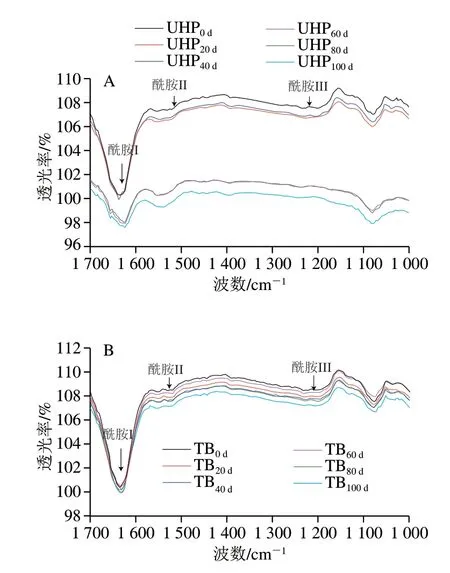
图 7 反复冻融过程中UHP(A)、TB(B)样品MP的FTIR变化Fig. 7 Changes in FTIR spectra of UHP (A) and TB (B) samples during repeated freeze-thaw cycles
蛋白质的肽键有几个特征振动带,包括酰胺I带(1600~1700 cm-1)、酰胺II带(1500~1600 cm-1)和酰胺III带(1200~1300 cm-1)[32],其中酰胺I和酰胺III带对于探讨蛋白质的二级结构十分有用[33],可定量分析获得各种二级结构的相对含量。酰胺I带主要来自C=O 的伸缩振动[34],酰胺III带来源于肽键的C—N伸缩振动和N—H面内振动。图7为不同冻融循环次数的UHP、TB样品MP的FTIR图,对酰胺I带(1700~1600 cm-1) 谱峰进行傅里叶自转积谱分析(半峰宽810,增强因子f=211),结合Origin软件里的Savitsk-Golay函数得到各个峰区对应的相对面积,即为各二级结构的相对含量[35],冻融过程中UHP、TB样品MP二级结构的相对含量见表1。
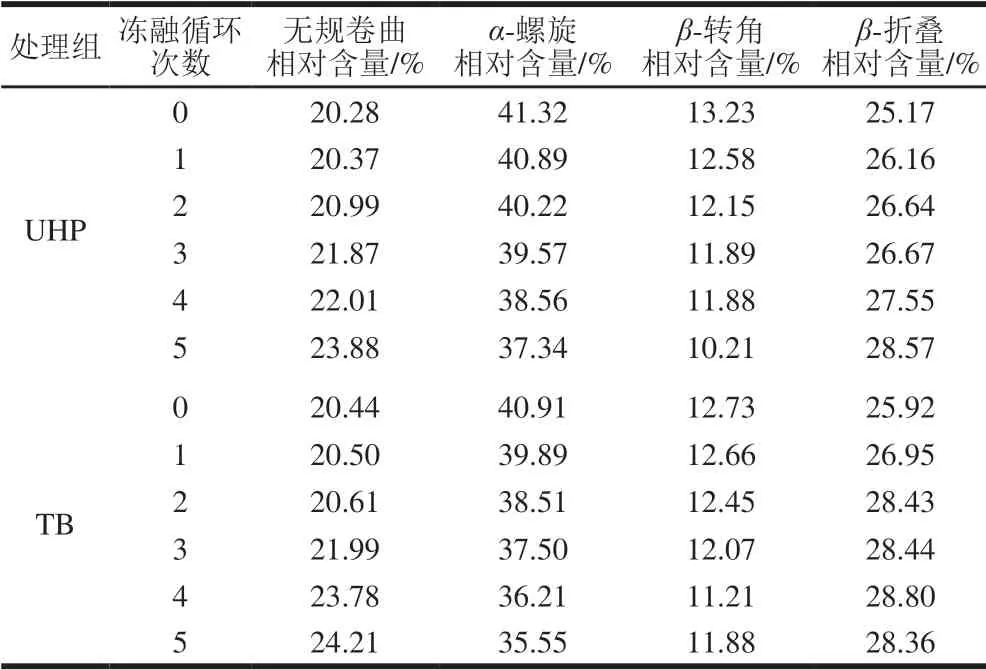
表 1 冻融过程中UHP、TB样品MP二级结构的相对含量Table 1 Relative contents of secondary structures in UHP and TB samples during freeze-thaw cycles
由表1可以看出,随着冻融循环次数的增加,UHP、TB样品MP的α-螺旋、β-转角相对含量降低,β-折叠、无规卷曲相对含量升高,并且UHP样品MP的α-螺旋相对含量下降趋势比TB缓慢。α-螺旋相对含量的减少表明鱼肉蛋白内部分子中的疏水基团暴露,表面疏水性增大。由于鱼肉水分在反复冻融过程中不断迁移,因此β-转角结构难以被维持,其相对含量出现较大幅度的下降,冻融5 次后,UHP、TB样品MP的β-转角相对含量分别将至10.21%和11.88%。在4 种二级结构中,β-转角相对含量最低,这可能是因为本实验在腌制鱼肉时添加了葡萄糖和白糖,小分子的糖类有助于MP形成片层结构,保持蛋白的分散结构[36]。与冻融0 次相比,冻融5 次后UHP样品的β-折叠相对含量增加量(3.40%)高于TB样品的β-折叠增加量(2.44%)。说明利用超高压和滚揉腌制加工的开背调味鱼在反复冻融过程中,尽管蛋白质会发生二级结构的变化,但鱼肉蛋白质仍能维持一定的稳定性,且超高压处理效果更优。
2.8 反复冻融过程中的UHP、TB样品MP的SDS-PAGE分析结果
由图8可以看出,UHP、TB样品MP的条带包括肌球蛋白重链(myosin heavy chain,MHC)、副肌球蛋白、肌动蛋白、原肌球蛋白和肌原蛋白,这些条带在电泳图中从上而下依次排列,与Sánchez-Alonso等[37]报道的结果一致。在冻融循环0~2 次,MHC、副肌球蛋白、肌动蛋白、原肌球蛋白、肌原蛋白条带变化并不明显;随着冻融循环次数的增加,尤其是冻融循环4~5 次时,MHC条带变细,肌动蛋白条带变淡,说明反复冻融使蛋白发生了降解。因为在加工和冻藏过程中MP中MHC比肌动蛋白和肌原蛋白更容易氧化[38], 从图8中也观察到MHC条带比其他条带变化较为明显,且TB组的MHC蛋白条带比UHP组样品明显变淡变细,这一结果与样品的总巯基和二硫键含量的变化结果一致,均是冻融循环次数对UHP组样品的 影响较小。尽管随着冻融循环次数增加,蛋白条带略有变化,但在冻融5 次时,仍能清晰可见5 条条带,说明超高压和滚揉腌制的开背调味鱼在反复冻融5 次后产品仍在可接受范围,没有发生明显的蛋白氧化现象。
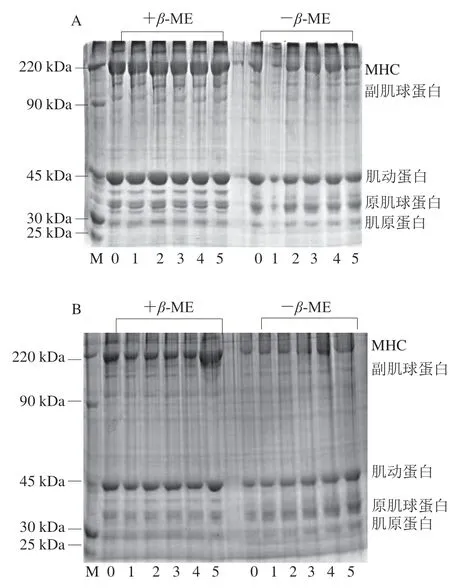
图 8 反复冻融过程中UHP(A)、TB(B)样品MP的SDS-PAGE变化Fig. 8 Changes in SDS-PAGE patterns of MPs in UHP (A) and TB (B) samples during repeated freeze-thaw cycles
2.9 反复冻融过程中的UHP、TB样品MP的微观结构变化

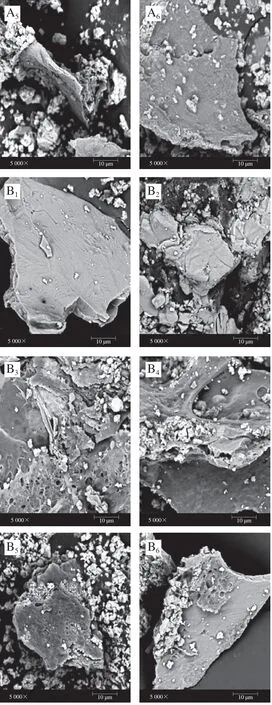
图 9 反复冻融过程中UHP、TB样品MP的微观结构变化Fig. 9 Microstructure changes of UHP and TB samples during repeated freeze-thaw cycles
冻融0 次时,UHP、TB样品MP冷冻干燥粉末表面平滑、结构致密、两者之间没有明显差异(图9A1、B1)。当反复冻融1 次时,UHP、TB样品MP表面相对平整,有微小褶皱,相比较而言,TB样品的褶皱比UHP样品更明显(图9A2、B2)。当反复冻融2 次时,UHP样品MP整体呈现块状结构,但出现细小的孔洞;而TB样品褶皱增加,表面微小的孔洞也增多,且呈蜂窝状,表面结构比UHP样品变得更粗糙(图9A3、B3)。
随着冻融循环次数的增加(3~5 次),UHP、TB样品MP表面的褶皱、孔洞变化均不明显,但TB样品褶皱程度均比UHP样品明显。这说明鱼肉经反复冻融循环,蛋白发生变性和氧化,会影响到MP的结构完整性,表现在MP表面会出现孔洞和褶皱。相比较而言,UHP样品比TB样品的结构破坏程度略轻,这与总巯基含量、二硫键含量、紫外光谱、傅里叶变换红外光谱等分析结果一致。
3 结 论
UHB、TB开背调味鱼在反复冻融过程(1~5 次)中品质变化如下:菌落总数始终维持在较低水平;随着冻融循环次数增加,TBARS值在冻融前3 次总体变化不显著(P>0.05),3 次后有显著升高趋势;TVB-N含量呈缓慢升高趋势,但冻融循环过程中始终低于淡水鱼鲜度TVB-N 20.00 mg/100 g的限量;冻融循环3~4 次后,样品的风味与新鲜开背调味鱼之间出现显著差异。
UHB、TB开背调味鱼在反复冻融过程(1~5 次)中MP特性变化如下:总巯基含量显著降低(P<0.05),二硫键含量显著升高(P<0.05);α-螺旋、β-转角相对含量降低,β-折叠、无规卷曲相对含量升高;MHC、肌动蛋白条带均有不同程度的变浅变细;扫描电子显微镜观察微观结构发现,冻融2 次时,样品表面明显出现孔洞、褶皱,表面结构粗糙。
对比UHP、TB样品发现,UHP样品的各指标变化趋势明显低于TB样品,说明UHP腌制方式可以延缓开背调味鱼在反复冻融过程的品质变化。
