平邑甜茶童区和成年区叶片基因表达差异
2021-05-24王新亮彭福田
王新亮,彭福田
(1 滨州学院学报编辑部,山东 256603)(2 山东农业大学园艺科学与工程学院,国家作物生物学重点实验室)
植物个体发育的过程分为童期和成年期。Zimmerman[1]指出:“童期是果树实生苗能花芽分化之前的一段时间,果树的童期一旦结束,就具有了开花的能力”。果树实生苗在空间上还存在一定的童区:以根茎为圆心,以童程(实生苗始果点至根茎的枝干长度)为半径的半球体空间[2]。童区不能形成花芽,而成年区能形成花芽。
关于童期向成年期转变的生理机制,学术界有3 种假说。“成花素假说”认为成花促进或抑制因子是一种简单的、特异的和广谱的激素,但是这种激素还没有被分离和确认[3];“营养分流假说”认为植株内部的源库关系能调控所有与阶段转变和成花有关的因子,成花诱导时,茎尖得到比非诱导条件时更好的同化物供应[4];“多因子控制假说”认为多种激素、代谢物、营养等物质参与成花诱导。遗传变异、过去和现在的生长状况对不同的种类或基因型或同样基因型在不同的环境下导致复杂的不同的限制因子[5]。近年来,研究者已经克隆到许多与植物阶段转变或成花有关的基因,越来越多证据支持“多因子控制假说”[6-9]。
为了进一步了解果树童区与成年区叶片在基因表达上的差异,本文以6 年生的平邑甜茶实生苗为试材,利用数字基因表达谱测序技术研究了在花芽分化期平邑甜茶童区与成年区叶片在基因表达上的差异,以期进一步揭示实生平邑甜茶由童区向成年区转变的分子机制,为分子育种提供理论基础。
1 材料与方法
1.1 材料
供试材料为6 年生平邑甜茶实生苗。花芽分化期(9 月5 日)分别取童区(Y2)和成年区(A2)的叶片储存于-80 ℃用于提取RNA。
1.2 RNA 提取和cDNA 制备
用TRIzol(Invitrogen)试剂盒提取平邑甜茶叶片中的总RNA,并通过Dynabeads Oligo(dT)(InvitrogenDynal)分离得到mRNA。用DEPC 水溶解RNA,并用1.0%的琼脂糖凝胶检测其质量。用Oligo(dT)作为引物合成cDNA 第一条链。
1.3 基因差异表达分析
基因差异表达分析由华大基因协助完成。反转录合成的双链cDNA 用NlaIII 限制性内切酶消化,并用磁珠纯化3′端cDNA 片段,然后在5′端接上Illumina adaptor1,再用MmeI 酶切CATG 位点下游17 bp 处,用磁珠去除3′片段,最后在3′端接上Illumina adaptor2,这样就得到了21 bp 标签库。经PCR 线性扩增15 个循环后,用6%TBE PAGE 纯化,然后通过Illumina 基因表达测序法测序。
1.4 原始数据处理
利用base calling 将测序得到的原始图像数据转化为序列数据,再经过特定处理转化为碱基序列标签。最后将所有标签对应到Genome Database for Rosaceae 网站苹果基因组V1.0(http://www.rosacea e.org/projects/apple_genome,Malus x domestica Geno me v1.0)[10]参考序列。
1.5 差异表达基因的筛选
差异表达基因的筛选标准:①不同处理间,同一基因的表达量存在2 倍及2 倍以上的差异;②错误率(false discovery rate FDR)小于0.001[11]。
2 结果与分析
2.1 标签概况
样品A2 产生了5 917 214 个原始序列,样品Y2 产生了6 126 204 个原始序列。去除杂质序列后,A2 产生了5 635 494 个标签,Y2 产生了5 825 279个标签。与苹果基因组预测基因(Apple Genome V1.0 predicted CDS)比对发现,A2 中有1 060 378个标签对应到16 284 个基因上,Y2 中有1 216 762个标签对应到15 719 个基因上(表1)。
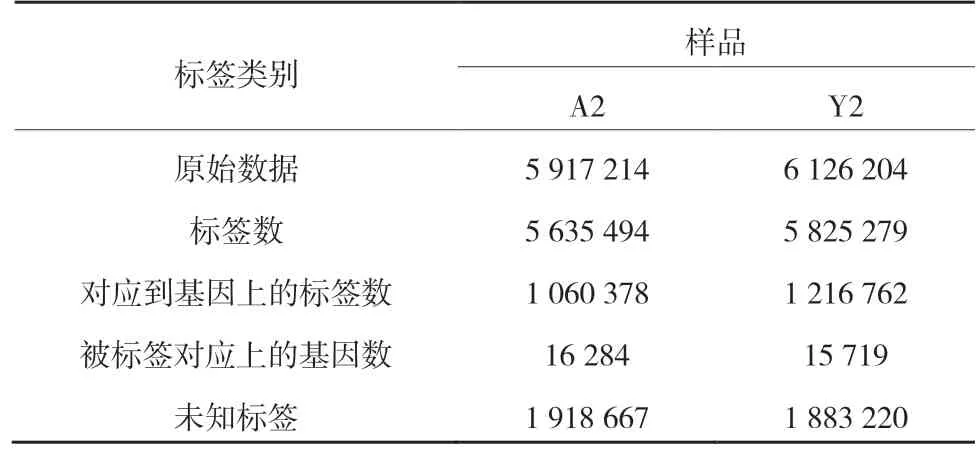
表1 每个样品获得标签数量
2.2 差异表达基因的确认与功能分析
通过与Y2 比较,在A2 中表达上调的基因有269 个,表达下调的基因有727 个。pathway 分析发现,差异表达基因中有606 个基因能对应到pathway中,排名前30 的已经在表2 中列出,差异表达基因主要集中在代谢途径、次生代谢物的生物合成、植物激素信号转导、植物-病原互作、核糖体、RNA转运、苯丙烷类生物合成、淀粉和蔗糖代谢这些途径中。
除了差异表达基因pathway 分析外,将差异比较大的基因(log2 Ratio(A2/Y2)≥7 or log2 Ratio(A2/Y2)≤-7)并通过NCBI 数据库的BLAST 工具与其他植物的基因序列进行比对。比对发现,表达上调的基因中主要参与含氮化合物转运与代谢,如:寡肽转运蛋白(MDP0000292270)和谷胱甘肽转移酶(MDP0000289011),生长素抑制蛋白(MD P0000323212),GTP 代谢相关基因(MDP0000617 187、MDP0000185787、MDP0000129000)。表达下调的基因涉及比较多的途径如脱落酸合成(MDP0000929213 ),E3 泛素蛋白连接酶(MDP0000770377)等。但是它们中的部分序列仍没找到明显相似的序列,这说明这些序列可能在其他植物中研究较少或是苹果中特有的(表3)。
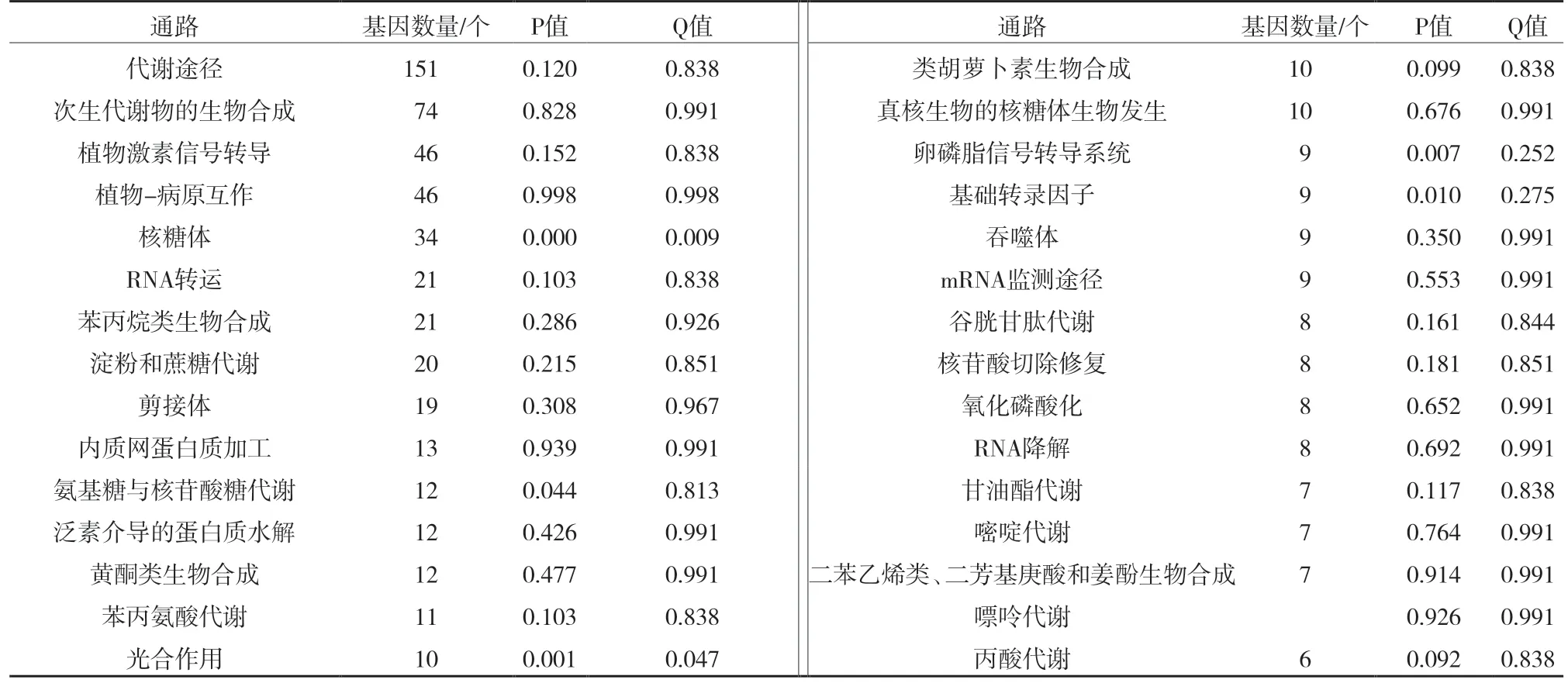
表2 差异表达基因前30位通路分析
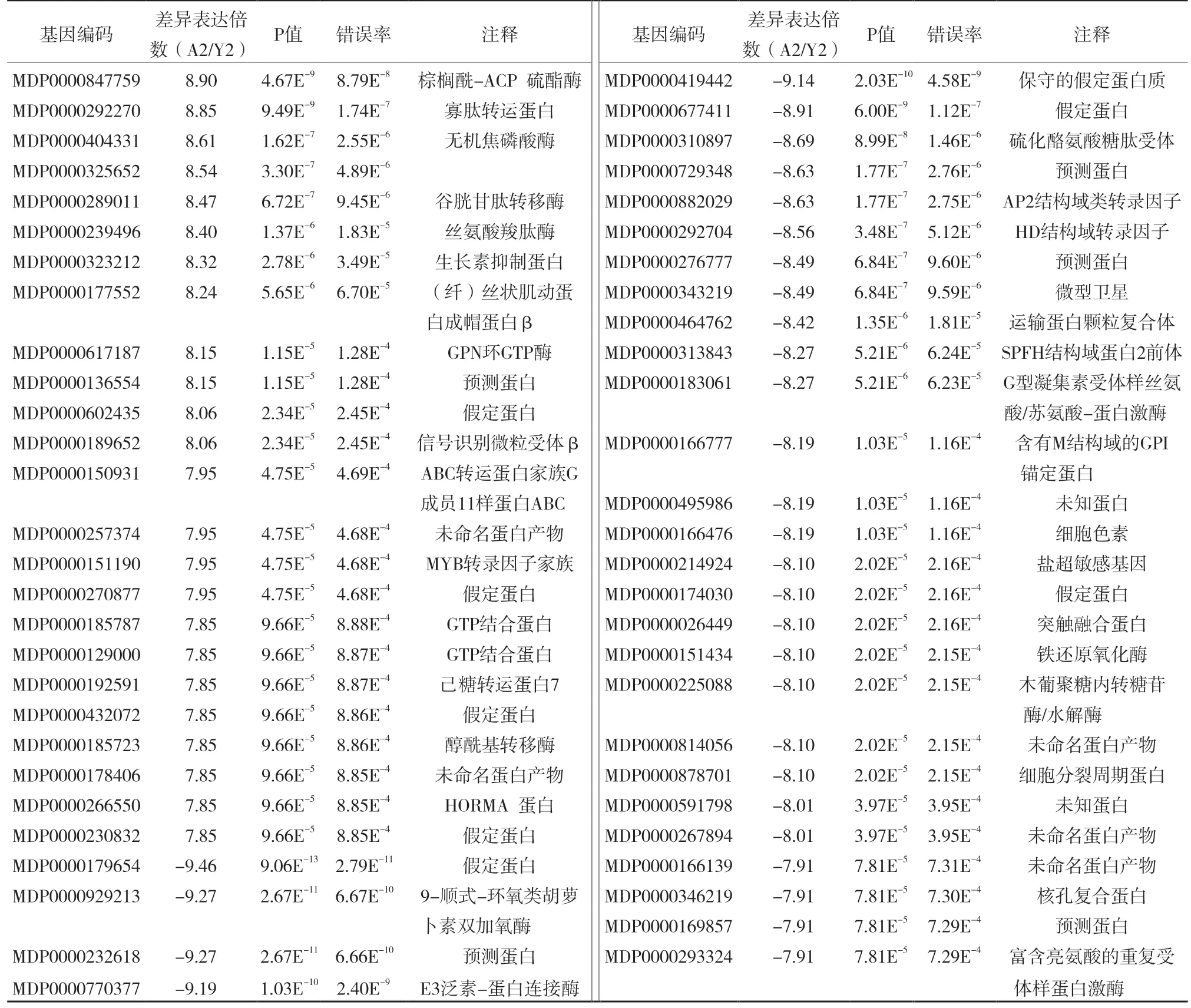
表3 表达差异比较大的基因
2.3 蔗糖和淀粉代谢酶编码基因在童区与成年区叶片中的表达差异
通过pathway 分析,在差异基因中查找到19 个参与蔗糖和淀粉代谢的基因,其中3 个在成年区表达量高,16 个在童区表达量高。淀粉合成关键酶AGPase(MDP0000182948)和淀粉酶(MDP00002 32760、MDP0000397284)编码基因在成年区的表达比童区低。蔗糖非发酵-1 型相关蛋白激酶 1(SnRK1)在植物糖信号转导和代谢途径中可能发挥着关键性开关的作用[12],我们在苹果预测基因中找到3 个SnRK1同源基因(MDP0000191788、MDP0000173500、MDP0000320932 ),其中MDP0000320932 在成年区的表达比童区高[Log2 ratio(A2/Y2)=1.46](表4)。
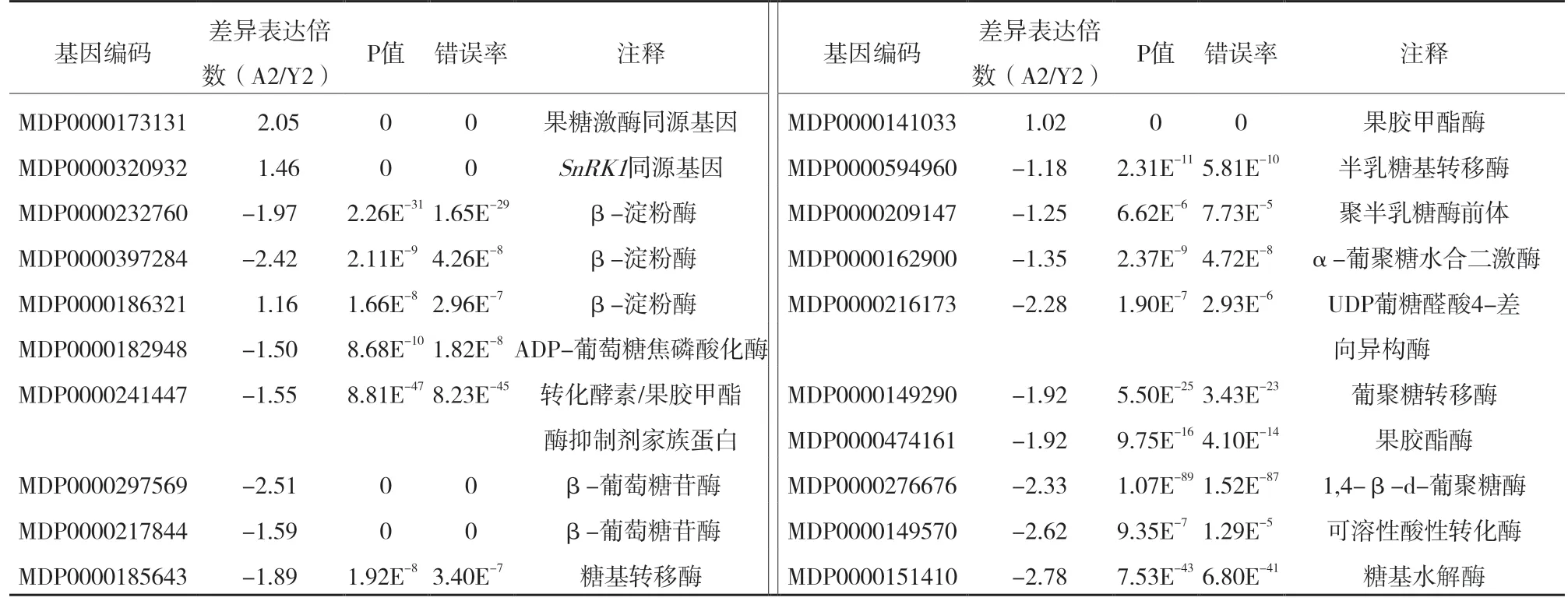
表4 参与蔗糖和淀粉代谢的差异表达基因
2.4 赤霉素和细胞分裂素合成关键酶编码基因的表达差异
赤霉素、生长素和细胞分裂素对花芽分化既有促进方面的报道,也有抑制方面的报道,不同类型赤霉素对苹果花芽分化的作用不同[13]。在本试验中,赤霉素合成主要酶编码基因(柯巴基焦磷酸合酶基因)MDP0000305417 和MDP0000147908 在成年区叶片中的表达高于童区,都是142 倍;1 个细胞分裂素合成关键酶—异戊烯基转移酶编码基因(IPT)在童区叶片中的表达是成年区的4.6 倍(表5)。

表5 参与赤霉素和细胞分裂素合成的差异表达基因
2.5 成花基因的表达差异
随着研究的不断深入,现在的研究主要集中到对成花基因的克隆及其功能的研究上。本试验中,4 个PAF1 复合体同源基因(MDP0000141365、MDP0000130422、MDP0000210032、MDP000014182 8)在童区叶片的表达分别是成年区叶片表达量的2.4、3.1、4.2、3.4 倍。1 个GI同源基因(MDP00001 71530)在成年区和童区叶片中的表达存在差异,其在童区叶片的表达量是成年区叶片的2.4 倍。本试验也发现1 个与PnFL-2同源性较高的基因(MDP0000322348)在成年区和童区叶片中的表达存在差异,其在童区叶片的表达是成年区叶片的13.2 倍。PFT1同源基因MDP0000220114 在童区叶片的表达是成年区叶片的6.4 倍。DNA 甲基化能终止春化作用,延迟开花[14]。本研究5 个甲基化相关酶基因中有4 个(MDP0000171530、MDP0000259 734、MDP0000312110、MDP0000223161)在成年区叶片的表达低于童区叶片(表6)。
3 讨 论
pathway 分析发现,差异基因中有606 个基因能对应到pathway 中,包括:碳和氮的固定、同化、代谢和降解,脂类合成和代谢,核酸代谢,次生代谢,植物激素信号等。因此,本研究也支持“多因子控制假说”。很多标签不能比对到预测基因上,这说明苹果基因组的预测基因可能并不完全。
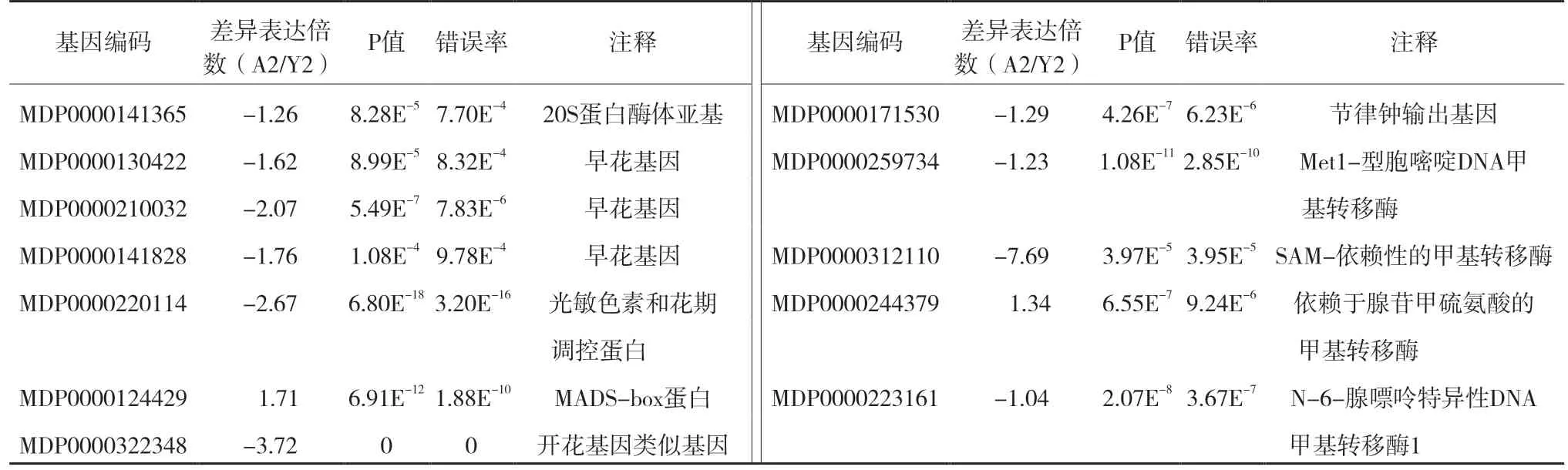
表6 成花基因的表达分析
3.1 花芽分化过程中糖代谢相关基因的表达
花诱导后,植物茎尖分生组织中的蔗糖浓度会激增[15],刺激细胞有丝分裂。苹果属植物施用葡萄糖、果糖或蔗糖后,能促进其成花。但甘露醇不能起到同样的作用,说明蔗糖在花诱导中是作为营养物质,而不是调节渗透性[16]。King 等[17]研究发现,蔗糖是助剂,而不是成花诱因。通过pathway 分析,在差异基因中查找到19 个参与蔗糖和淀粉代谢的基因,其中3 个在成年区表达量高,16 个在童区表达量高。淀粉动员是花诱导早期的重要步骤[18]。淀粉合成关键酶AGPase(MDP0000182948)和淀粉酶(MDP0000232760、MDP0000397284)编码基因在成年区的表达比童区低,说明可能淀粉合成和降解均降低。这时光合产物可能主要被运往花芽,为来年开花积累能量。SnRK1 在植物糖信号转导和代谢途径中可能发挥着关键性开关的作用[12],我们在苹果预测基因中找到 3 个SnRK1同源基因(MDP0000191788、MDP0000173500、MDP00003209 32),其中MDP0000320932 在成年区的表达比童区高,这表明SnRK1 也可能在成花诱导方面起一定的作用。
3.2 花芽分化过程中激素合成相关基因的表达
研究证明,赤霉素能促进拟南芥开花[19]。而不同类型赤霉素对苹果树花芽分化的作用不同[13]。超表达GA-20 氧化酶基因后,植物开花提前[20]。在本试验中,赤霉素合成主要酶编码基因(MDP0000305417 和MDP0000147908)在成年区叶片中的表达高于童区。说明苹果可能通过赤霉素上调LEAFY的表达来促进花器官的发育[21]。1 个IPT基因(MDP0000295823)在成年区叶片中的表达低于童区。IPT 是植物异戊二烯类细胞分裂素[反式玉米素(tZ),二氢玉米素(DHZ),顺式玉米素(cZ),异戊烯基核糖基腺嘌呤(iP)]合成的关键限速酶[22]。李广等[23]研究发现,在花芽分化后期侧柏叶片中的玉米素含量是降低的。本研究发现,成年区叶片中IPT表达量低于童区,可能是导致细胞分裂素含量降低的直接原因。
3.3 花芽分化过程中成花基因表达差异
在拟南芥中,FRI、FRL1、PAF1 和FLC 抑制花芽分化,而SOC1、FT、AP1 和LEAFY 促进花芽分化[21,24]。本试验中,4 个PAF1 复合体同源基因(MD P0000141365、MDP0000130422、MDP0000210032、MDP0000141828)在成年区叶片的表达低于童区叶片。PAF1 复合体刺激FLC的表达来抑制AP1和LEAFY的表达,延迟成花,因此平邑甜茶成年区秋季可能主要靠下调抑制花芽分化的PAF1 复合体同源基因的表达的方式来上调AP1同源基因(MDP0000124429)最终达到花芽分化。另外,GI参与光信号生物钟响应。OsGI超表达造成水稻延迟开花,而通过RNAi 沉默OsGI后的水稻开花提前[25]。但是拟南芥中GI表现出相反的功能[26]。本试验也发现1 个GI同源基因(MDP0000171530)在成年区和童区叶片中的表达存在差异,其在成年区叶片的表达低于童区叶片,这说明它的功能可能和水稻OsGI一样。Kim 等[27]报道,牵牛花PnFL-2受长日照诱导,其转基因拟南芥在长日照条件下开花提前,PnFL-2 属于TIFY 转录因子家族的ZML 亚族。本试验也发现1 个与PnFL-2同源性较高的基因(MDP0000322348)在成年区叶片的表达低于童区叶片。据李肖琴[28]报道,MDP0000322348 虽然也属于TIFY 转录因子家族,但其归属于TIFY 亚族,所以可能MDP0000322348 的表达特性和功能与牵牛花PnFL-2的不同。拟南芥光敏色素开花时间蛋白1(phytochrome and flowering time protein 1,PFT1)作用于光敏色素B 的下游调节FT的表达[29]。任保兰等[30]发现,红掌Aa PFT1在童期表达量高于成年期,而本研究发现,PFT1同源基因MDP0000220114成年区叶片的表达低于童区叶片。DNA 甲基化能终止春化作用,延迟开花[14]。齐思言[31]研究发现,苹果基本不成花的腋芽基因DNA 甲基化水平明显高于易成花的短枝顶芽。菊花叶片经短日照处理后DNA 甲基化也显著降低[32]。石玉波等[33]认为,DNA甲基化降低与花芽分化之间具有一定的相关性。本研究发现4 个甲基化相关酶基因在成年区叶片的表达低于童区叶片,这说明甲基化也可能在调节平邑甜茶成年区花芽分化过程中起重要作用。
