靶向共价闭合环状DNA抗HBV治疗的研究进展
2021-05-17王艺颖刘熙称朴荣利秦俊杰
王艺颖,刘熙称,朴荣利,秦俊杰
吉林大学第一医院 肝病科,长春 130000
HBV感染已成为全球严重卫生问题,全球慢性乙型肝炎(CHB)患者已达2.57亿,其显著增加肝硬化、肝衰竭和肝细胞癌的罹患风险。2017年全球肝炎报告显示,每年约有65万人死于HBV感染,给人们乃至社会带来了沉重负担[1]。1965年由Blumberg等[2]发现的HBV,随后被证实为一种具有内源性DNA聚合酶活性的DNA病毒。1982年,Summers等[3]研究发现HBV聚合酶将RNA中间体逆转录为不完整且松弛的环状DNA(relaxed circular DNA,rcDNA),这一环状DNA随后转化为共价闭合环状DNA(cccDNA),形成病毒转录的模板,并居于宿主细胞核中。这些突破性的发现开创了HBV分子生物学时代,重组HBV疫苗的开发随之拉开序幕。据报道[1,4],世界范围内的预防性疫苗接种计划已明显降低5岁以下儿童的HBV患病率。当前用于HBV感染的标准疗法包括聚乙二醇化干扰素α和核苷酸类药物(NAs)[5]。自1998年以来,已经批准了6种NAs应用于临床实践中,包括拉米夫定、恩替卡韦、替比夫定、阿德福韦酯及替诺福韦[6],但所面临的巨大考验也不容忽视:首先,IFNα对cccDNA转录的长期和可持续抑制具有重要作用,但干扰素的应答率欠佳;其次,NAs可以通过靶向病毒RNA依赖性DNA聚合酶来抑制HBV复制,但对现有的cccDNA微染色体没有直接影响,NAs终止后常见病毒复发,甚至在达到治疗终点的患者中亦如此[7]。尽管使用IFNα和NAs已观察到有效的抗病毒应答,但目前仍无法完全清除cccDNA,为彻底治愈HBV感染,迫切需要根除肝内cccDNA的新治疗策略[8-10]。本文对关于cccDNA的当前知识及其作为治疗靶标的潜在作用作一综述,旨在探索消除病毒cccDNA潜在治疗策略的前景及曙光。
1 HBV的身世之谜
1.1 HBV的生命周期——cccDNA的持续性 肝细胞是HBV易感染的靶细胞之一,HBV通过血液到达肝脏感染肝细胞。HBV与位于宿主肝细胞上的硫酸肝素蛋白聚糖相互作用,牛磺胆酸钠共转运多肽(sodium taurocholate cotransporting polypeptide,NTCP)与之结合进入肝细胞[6](图1步骤-1),随后带有rcDNA的核衣壳(图1步骤-2)被释放到细胞质中并进入细胞核,进入核内的rcDNA利用宿主DNA修复机制转化为高度稳定的游离DNA,即cccDNA[11-12](图1步骤-3)。cccDNA被转录为HBV前基因组RNA(pregenomicRNA,pgRNA)和几个亚基因组RNA[13](图1步骤-4)。HBV DNA整合感染肝细胞后立即进入宿主基因组[14]。完成转录和翻译后(图1步骤-5),pgRNA被包装并组装在核衣壳中(图1步骤-6),并被转录成松弛的环状DNA(图1步骤-7),最后成熟的衣壳,即含有rcDNA的衣壳,可以通过细胞分泌途径,并以包膜和感染性病毒体的形式释放[15]。重要的是,细胞内成熟衣壳中新形成的后代rcDNA(图1步骤-8),如来自传入病毒体的rcDNA,也可以被递送至宿主细胞核并启动额外cccDNA分子的形成,从而有助于病毒的持久性[7,16]。

图1 HBV复制周期示意图[4,17-18]
1.2 HBV的基因组信息 HBV基因组呈环状结构,含内部核衣壳核心的外部脂蛋白包裹部分双链的DNA[16]。 HBV DNA长度仅为3.2 kb,包含4个重叠的开放阅读框,分别为S、C、P和X。cccDNA缠绕在组蛋白上形成病毒微染色体,是所有病毒RNA转录物(pg、preS、S和X)编码七种蛋白质的独特模板(图1步骤-5):pgRNA充当病毒聚合酶介导的逆转录和rcDNA后续合成的模板;HBcAg是另一种基因产物核心蛋白,它参与病毒核衣壳的构成;此外前核心蛋白(HBeAg)是一种蛋白水解加工的病毒蛋白。HBsAg是所有不同长度的表面蛋白的统称,其中包括病毒表面蛋白S、M和L(其中S从S mRNA进行翻译,M从preS2+ S mRNA进行翻译,L从preS1+PreS2+S mRNA进行翻译)。最后,cccDNA还编码一种非结构蛋白即病毒蛋白X(HBV regulatory X,HBx),可能起转录反式激活因子的作用,并在调节病毒基因表达中发挥作用。
据研究发现,HBV DNA也可整合到宿主染色体中。病毒DNA的整合似乎在病毒复制的过程中并没有发挥直接作用,但有报道[14]发现,HBV DNA整合可能通过调节基因表达使细胞环境更易于病毒复制,可能在肝细胞癌变中起重要作用。目前有关cccDNA的大多数知识都来自于鸭DHBV模型,cccDNA研究的主要障碍之一是缺乏可靠的、可量化的体外培养系统,该系统无法生成真正的HBV cccDNA。有研究[19]发现HepG2-HBV/loxP细胞系统在研究cccDNA相关生物学机制和开发新型cccDNA靶向药物方面具有强大的潜力。
2 消除cccDNA的新策略
目前使用NAs的抗病毒疗法靶向病毒聚合酶并抑制逆转录。它们阻止病毒基因组rcDNA的合成,以防止成熟衣壳的形成和传染性子代病毒的释放,从而通过细胞内cccDNA扩增来减少cccDNA的补充。但是,NAs不能靶向已建立的cccDNA,因此不能治愈慢性HBV感染。
2.1 基于DNA损伤修复机制的设想 宿主细胞DNA损伤修复机制将rcDNA识别为损伤的DNA,并介导其转化为cccDNA[20]。近年来有报道[21]称,酪氨酰DNA磷酸二酯酶2(Tyrosyl DNA phosphodiesterase 2 )及FEN1(flap structure-specific endonuclease 1),在cccDNA形成的过程中扮演重要角色,除了进行实际的DNA修复反应以从rcDNA形成HBV cccDNA的酶促机制外,其他包括调节HBc磷酸化状态以及衣壳核导入和脱壳的宿主蛋白激酶与磷酸酶等宿主因素,可能也参与控制cccDNA的形成。但是,由于迄今确定的与cccDNA形成有关的宿主因子也与许多基本细胞过程有关,考虑到潜在的严重副作用,它们似乎不太可能作为治疗靶标[7]。
在细胞核中,与cccDNA微型染色体结合的HBx蛋白可在表观遗传上发挥其影响,同时启动并维持cccDNA的转录。HBx参与DNA修复途径,宿主蛋白损伤特异性DNA结合蛋白1(damage-specific DNA-binding protein 1,DDB1)与其结合,对于病毒复制尤为重要[22],通过募集含有DDB1的E3泛素连接酶以降解宿主限制因子SMC5/6(structural maintenance of chromosomes 5/6),以允许cccDNA进行HBV转录,否则将被转录抑制[5]。鉴于其在HBV生命周期中的重要作用,除了影响表观遗传转录并刺激众多细胞转导途径外,靶向HBx成为一种可行的治疗策略。硝唑尼特[2-乙酰氧基-N-(5-硝基-2-噻唑基)苯甲酰胺]是一种口服的噻唑类抗感染药。Sekiba等[23]通过化合物筛选用于药物重新定位的方法发现硝唑尼特显著抑制HBx-DDB1相互作用,从而抑制了病毒转录、蛋白质表达和cccDNA水平,为HBV提供新的治疗选择。
2.2 通过基因编辑技术沉默cccDNA表达 锌指核酸酶(zinc fingernucleases,ZFN)、转录激活子样效应子核酸酶(transcription activator-like effector nucleases,TALEN)和成簇的规则间隔的短回文重复序列/CRISPR相关(clustered regularly interspaced short palindromic repeats/CRISPR-associated Cas9 nucleases,CRISPR/Cas9)系统等基因组编辑技术可以有效地破坏HBV cccDNA[24-27]。DNA将来自四种不同物种的CRISPR/Cas9系统与靶向HBV基因组保守区域的指导RNA在细胞系中共表达,发现源于化脓性链球菌(streptococcus pyogenes,Sp)和嗜热链球菌(streptococcus thermophilus,St)的CRISPR/Cas9系统通过靶向HBV基因组的保守区域,从而阻断了HBV复制,同时超过90%的HBV cccDNA转染6 d后被降解。研究发现HBV cccDNA甲基化影响CRISPR/Cas9的抗病毒活性,而单核苷酸HBV遗传变异不会影响St CRISPR/Cas9的抗HBV活性,表明其可用于靶向许多HBV变异。而两个或多个错配会破坏或阻断CRISPR/Cas9活性,使其宿主DNA不太可能成为靶向。最重要的是,深度测序表明Sp CRISPR/Cas9引起脱靶诱变,而St CRISPR/Cas9对宿主基因组没有影响,St CRISPR/Cas9系统代表了具有高抗HBV活性的最安全的系统[28]。
ZFN作为一种DNA核酸内切酶,具有基因组编辑潜能,可用于在特定的靶位点介导DNA双链发生断裂反应,并在裂解位点处产生序列改变而发挥修复作用[24]。Weber等[29]设计了3种靶向HBV聚合酶,基因X和核心的ZFN,并分别通过自身互补的腺相关病毒载体将其分别导入HepAD38细胞,实验中发现这些针对HBV的ZFN可以有效抑制HBV主动复制,并抑制HBV持久性的细胞模板。TALEN与ZFN相似,但TALEN的DNA结合结构域是高度重复的,并且源自转录激活因子样III效应子。2013年发现TALEN可以降低细胞培养物中HBV产生的效率[30]。随后研究人员使用TALEN介导的同源性定向重组将人工初级miRNA引入HBV基因组可以提高TALEN的抗病毒效力[31]。
2.3 通过表观遗传修饰沉默cccDNA转录 表观遗传学研究的重点是基因表达的调控机制,包括DNA甲基化、组蛋白修饰、染色质重塑和非编码RNA干扰等。HBV cccDNA以组蛋白修饰的微染色体的多个拷贝形式存在于细胞核中,因此所有这些调节机制都会影响HBV复制和持续感染[6]。IFNα通过募集组蛋白脱乙酰基酶1(histone deacetylase 1,HDAC1)减少与cccDNA结合的乙酰化组蛋白H3来降低HBV cccDNA转录活性[32]。shRNA(short hairpin RNA)可以诱导HBV cccDNA甲基化,从而抑制其在人肝癌细胞中的转录[33]。NIRF(Np95/ICBP90-like RING finger protei)一种新型的E3泛素连接酶,可使转染了pAAV-HBV1.3的HepG2细胞中的HBV DNA复制,HBsAg和HBeAg的分泌受到抑制作用。此外,NIRF还能对表达HBV的小鼠模型中HBV的复制和分泌起到抑制作用。NIRF减少HBV cccDNA结合的H3组蛋白的乙酰化。这些结果表明,不仅可以通过与HBc直接相互作用抑制HBV复制,还能减少与H3组蛋白的乙酰化[34]。姜黄素可通过下调cccDNA结合的组蛋白乙酰化来抑制HBV复制[35]。沉默类视黄醇X受体α会导致cccDNA形成和病毒抗原产生升高,而它的激活会降低HBV感染效率[36]。DNA甲基化和组蛋白乙酰化是cccDNA形成所必需的,这些共同的发现表明,对其调控可使cccDNA转录失活,从而抑制HBV复制[6]。Yuan等[37]的研究进一步表明组蛋白脱乙酰基酶11特异性降低了cccDNA结合的组蛋白H3的乙酰化水平,通过cccDNA转录的表观遗传抑制作用来阻断HBV复制,为抗HBV药物的开发提供了重要参考。
3 未来展望
经过几代人的努力,在HBV的各个方面都取得了不容忽视的进步,不仅发现了可以有效抑制病毒复制的预防性疫苗和抗病毒药物,并且几种新颖的抗HBV治疗药物正在临床前开发或早期临床试验中[7,38](表1)。但完全消除核cccDNA仍遥遥无期,主要的努力仍然是致力于开发能够清除或沉默HBV cccDNA的有限疗法[39]。基因编辑的进展,特别是CRISPR-Cas9技术,这是唯一已知能破坏先前存在的cccDNA,但脱靶和体内递送方法等问题仍面临着巨大考验。尽管基于基因编辑和基于表观遗传修饰的策略都需要克服挑战,并且从基础研究到临床应用还有很长的路要走,但却已显示出光明的前景。
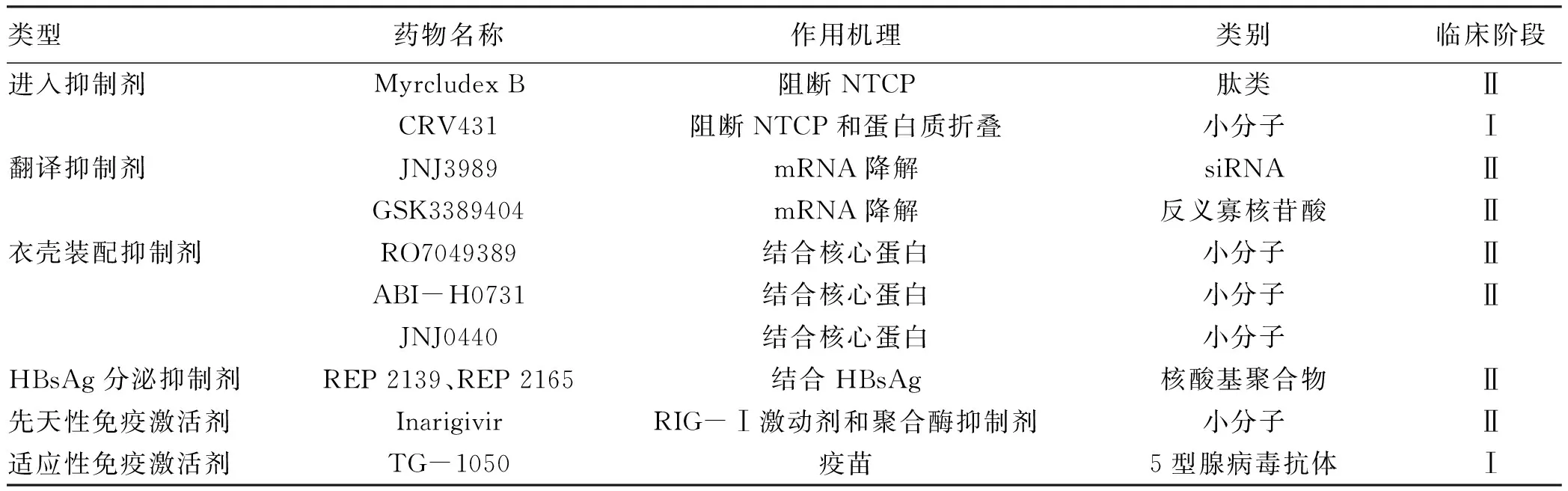
表1 新研发的抗HBV感染的代表药物
利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:王艺颖负责资料分析、撰写论文;刘熙称负责收集数据,修改论文;朴荣利负责课题设计、拟定写作思路;秦俊杰负责撰写文章并最后定稿。
