中国传统发酵食品来源典型小分子脂肪酸酯合成微生物及酶的研究进展
2021-05-15王晓程徐友强李秀婷孙宝国
王晓程,徐友强,李秀婷,孙宝国
(北京工商大学 北京食品营养与人类健康高精尖创新中心 北京市食品添加剂工程中心 北京100048)
中国传统发酵食品的历史悠久,风味独特,品种多样,深受广大人民喜爱,是食品工业的重要组成部分。酯类物质是传统发酵食品中的一类重要风味物质,以小分子脂肪酸酯类为代表。传统发酵食品来源的小分子脂肪酸酯是一类低分子质量的具有芳香气味的化合物。以乙酸乙酯、丁酸乙酯、戊酸乙酯、己酸乙酯和辛酸乙酯等为典型代表,呈现独特的风味,例如:乙酸乙酯具有苹果样风味,味短,刺激带涩;丁酸乙酯略带脂肪臭气味,似菠萝样果香气味,味涩,爽口;己酸乙酯有菠萝样果香气味,味甜爽口,带刺激涩感[1]。小分子脂肪酸酯本身的特性,使其成为决定发酵食品感官和风味的一类重要化合物。
发酵食品中的小分子脂肪酸酯类主要来源于微生物的合成代谢。研究表明,发酵食品制作过程中有大量微生物参与,微生物可以分泌各种酶,包括酯酶、脂肪酶、蛋白酶、半纤维素酶、纤维素酶和淀粉酶等,作用于原料中的淀粉、蛋白质和脂肪酸等。在一系列酶的作用下,一方面产生有利于微生物生长的小分子糖类和氨基酸等主要碳源和氮源,另一方面产生益生元,如低聚糖以及功能性短肽等物质,提升了发酵食物的营养价值。在发酵过程中,微生物生长代谢产生的有机酸和醇,可在酯酶和脂肪酶等羧酸酯水解酶催化下合成小分子脂肪酸酯,增强发酵食品的独特风味[2]。这些小分子脂肪酸酯,如乙酸乙酯、丁酸乙酯、己酸乙酯,在人体内水解产生己酸、丁酸和乙酸等短链脂肪酸,对人体健康具有重要影响。如丁酸可以修复肠黏膜,治疗肠炎以及抑制多种癌细胞的增值,乙酸可以消除疲劳,抗衰老和增强肾脏功能等。研究解析发酵食品中小分子脂肪酸酯的合成机制,对于确保发酵食品的产品品质具有重要意义。
微生物的酯合成能力来源于其所表达的酯合成酶。微生物中具有催化酯合成能力的酶有很多种,本文主要介绍酯酶(EC 3.1.1.1)和脂肪酶(EC 3.1.1.3),二者催化反应的最适温度和最适pH 值随底物及发酵食品中酶浓度的变化,以及在食品发酵过程中催化小分子脂肪酸酯的合成情况。
1 酯合成微生物的多样性
中国传统发酵食品品类丰富,以发酵酒、食醋、酱油、泡菜、发酵乳制品为典型代表,均具有“多微共窖”的特点。在发酵过程中,部分微生物对小分子脂肪酸酯类化合物的形成起到了重要作用,如文献报道(见表1),多种具有酯合成的微生物,包括细菌、酵母菌和霉菌均被发现于发酵食品中。
1.1 酯合成细菌的多样性
传统发酵食品营养丰富、风味独特,这些风味物质的来源主要包括3 个方面,原料的带入、发酵过程中的化学转化以及生物作用。其中,生物作用即微生物代谢产生了大量的风味物质。细菌对于小分子脂肪酸酯类风味物质的合成具有重要作用。例如:一些重要的产脂肪酶的细菌可以通过深层发酵方式产生胞外脂肪酶,包括芽孢杆菌、假单胞菌、伯克霍尔德氏菌等[28];微生物酯酶在细菌中亦有广泛分布,在链球菌、葡萄球菌、新型隐球菌、乳酸菌、魏斯氏菌等多种细菌中均有发现[29]。
我国传统发酵食品领域尤其重视具有小分子脂肪酸酯等风味物质合成功能的细菌的研究与应用[30]。例如:在中国传统白酒发酵领域,芽孢杆菌是中国主流大曲白酒发酵中对风味起重要作用的细菌[31],杨帆等[32]研究了对茅台大曲风味有突出贡献的3 株芽孢杆菌:地衣芽孢杆菌MTDB-01、地衣芽孢杆菌MTDB-02、枯草芽孢杆菌MTDB-03,根据微生物丰度解析确定3 株芽孢杆菌分别约占大曲中细菌总数的28%,37%,23%,由此微生物组群形成了较为稳定的细菌发酵系统;梁龙元[33]筛选获得高产酯酶的芽孢杆菌,其酯酶催化底物具有选择性,对己酸的亲和及催化酯化效率较高,可以有效提高白酒中己酸乙酯的含量;郭通航等[34]从变质食用油中分离出1 株具有己酸乙酯合成能力的伯克霍尔德氏菌X10,该菌株在优化发酵条件下可产生胞外酯酶,用于提高浓香型白酒的风味和品质;黄丹等[35]从泸香型大曲中成功筛选出1株血红鞘氨醇单胞菌,该菌在以黄豆粉为氮源,玉米粉为碳源,pH 7.0,32 ℃,150 r/min 条件下恒温培养48 h,其酯化酶的酶活可达18.26 U/mL。
食醋是一类重要的调味品,不仅是我国的一类代表性传统发酵食品,在世界范围内也广泛分布,其产香的小分子脂肪酸酯以乙酸乙酯为代表,催化这种酯合成的绝对优势细菌包括乳杆菌以及醋酸杆菌[12]。聂志强等[36]采用宏基因组学的方法分析了天津独流老醋醋酸发酵阶段的细菌组成,发现乳杆菌和醋酸杆菌的丰度较高。乳杆菌是一类重要的益生菌,其主要功能是产生大量乳酸并且合成酯化酶催化乳酸乙酯的生成,形成食醋的独特风味[14]。雷玛莎[37]在研究微生物及其酶在固态发酵制醋中的作用时发现,乳酸菌的发酵有一个重要的生化过程,即可以使最后生成的部分乙酸与乙醇生成乙酸乙酯,对食醋独特的浓郁香味具有重要的感官贡献。醋酸杆菌是食醋中一类重要的生产菌株[14],王宗敏[38]研究发现醋酸菌是与食醋中风味物质合成高度相关的核心微生物菌群之一,从镇江香醋醋醅中筛选到1 株巴斯德醋酸菌(Pasteurella acetate),可产生酯酶催化醋酸与乙醇合成乙酸乙酯。王宗敏等[39]还从镇江香醋中分离得到环状芽孢杆菌(Bacillus circulans),该菌株可以耐受6%的乙醇,在pH>4、醋醅浸取液的比例在20%~40%时生长良好,并可促进小分子脂肪酸酯等挥发性物质的生成。
传统发酵食品的典型代表还包括酱油、泡菜和乳制品等,其中丰富的细菌可以产生多种风味化合物。在酱油研究中,Xiao 等[21]研究微生物菌群与酱油风味的关系,结果表明细菌对风味的形成贡献比真菌更突出,其中乳杆菌(植物乳杆菌[40-41])、明串珠菌、无色杆菌和片球菌与风味密切相关;Harada 等[42]使用气相色谱-质谱联用的方法研究了乳酸菌和酵母菌代谢产物对酱油香气特征的影响,确定了微生物源重要香气成分:乙酸乙酯;刘卓[43]研究了耐盐乳酸菌对发酵工艺和酱油主要风味的影响,结果表明耐盐乳酸菌的添加可以促进酵母菌的生长,且对中后期酱油主体风味的形成和提高具有良好效果,香气成分的检测表明,乙酸乙酯等11 种香气成分明显提高,说明耐盐乳酸菌是改善酱油风味的重要细菌资源。魏斯氏菌(Weissella sp.)是酱油发酵过程中的优势细菌,可以代谢合成乙酸、丙酸、丁酸、戊酸等短链脂肪酸;李巧玉[44]从高稀态酱油中筛选到3 株魏斯氏菌:类肠膜魏斯氏菌(Weissella enterica)、融合魏斯氏菌(Weissella faecalis)和食窦魏斯氏菌(Weissella sinensis),并发现类肠膜魏斯氏菌是酱醪来源魏斯氏菌中具有产酯酶和类酯酶功能的菌株,其存在提升了成品酱油的风味。在泡菜研究中,朱琳等[45]采用高通量测序分析了萝卜泡菜中细菌的多样性,其中优势菌群且具有酯合成性能的细菌包括乳酸菌、魏斯氏菌和片球菌;魏晓明等[23]在黄秋葵泡菜研制中,添加4%植物乳杆菌、0.3%生香酵母,在此条件下泡菜的酸味和酯香味协调,感官评分最高;张基圣等[46]以低盐泡菜为研究对象,其发酵前期的主要优势细菌是乳酸球菌和食窦魏斯氏菌,发酵中、后期的主要优势菌群是戊糖乳杆菌,乳酸球菌是乙酸乙酯合成的主要功能菌。在发酵乳制品研究中,李伟程等[47]在解析传统发酵乳制品中微生物的多样性时,发现酸牛奶样品中细菌属以乳球菌属为主,酸马奶和酸驼奶样品中优势细菌以乳杆菌属为主,经HS-SPME/GC-MS[48]检测方法确定这些细菌是产C6~C8[49]小分子脂肪酸酯的主要菌种;王小标[50]在酸马奶乳酸菌与酵母菌互生关系的研究中,发现干酪乳杆菌MLS5 单菌发酵酸马奶中含有32 种风味物质,其中酯类物质有7 种,包括乙酸乙酯、己酸乙酯、辛酸乙酯、癸酸乙酯等。

[3-27]物生微的力能成合酯酸肪脂子分小有具中品食酵发统传1表[3-27]Microorganisms with small molecule fatty acid ester synthesis ability in traditional fermented foods Table 1 菌霉菌母酵菌细(Rhizopus霉(Aspergillus niger)、根霉曲黑极(Saccharomyces cerevisiae)、南母酵酒酿菌杆孢(Burkholderia sp.)、芽菌氏德尔霍克伯、(As-霉曲色灰(Mucor sp.)、烟霉sp.)、毛酵赤毕常(Candida antarctica)、异母酵丝假孢芽状(Bacterium sp.)、梭菌杆(Bacillus sp.)、拟(Monascus霉曲红色pergillus fumigatus)、紫(Torulosis母酵拟(Pichia anomala)、球母(Staphylo-菌球萄(Bacillus fusiformis)、葡菌杆(Monascus ruber)霉曲红色purpureus)、红sp.)(Sphingomonas菌胞单醇氨鞘红coccus sp.)、血)sanguinis霉头)、犁(Monascus purpureus霉曲红色紫赤)、毕(Saccharomyces cerevisiae母酵酒酿(Staphylococ-菌球萄)、葡(Bacillus sp.菌杆孢芽、(Peni-霉)、青(Mucor sp.霉)、毛(Absidia sp.(Saccha-母酵孢膜复囊)、扣(Pichia sp.母酵属菌)、泛(Enterobacter sp.属菌杆)、肠cus sp.)、(Aspergillus fumigatus霉曲)、烟cillium sp.母酵姆汉科威常)、异romycopsis fibuligera属菌)、梭(Weissella sp.菌氏斯)、魏(Sodalis sp.)(Rhizopus oryzae霉根米母酵香)、酒(Wickerhamomyces anomalus)(Lactobacillus sp.菌杆酸)乳(Clostridium sp.)(Brettanomyces sp.)、根(Aspergillus sp.霉)、曲(Mucor sp.霉毛(Saccharomycopsis fibulig-母酵孢膜复囊扣(Acineto-菌杆动)、不(Lactobacillus sp.菌杆乳、)、犁(Penicillium sp.霉)、青(Rhizopus sp.霉(Wickerhamomyces母酵姆汉克威常)、异era杆孢)、芽(Pseudomonas sp.菌胞单)、假bacter sp.)(Absidia sp.酶头母酵丝)、假(Pichia sp.母酵赤)、毕anomalus)、链(Acetobacter sp.菌杆酸)、醋(Bacillus sp.菌)、裂(Hansenula sp.母酵逊)、汉(Candida sp.(Actinomadura菌线)、放(Streptococcus sp.菌球母酵拟)、球(Schizosaccharomyces sp.母酵殖)sp.)(Torulopsis sp.(As-霉曲)、黑(Aspergillus oryzae霉曲米酵氏)、鲁(Candida versatilis母酵丝假变多(Staphylococ-菌球萄)、葡(Bacillus sp.菌杆孢芽)pergillus niger(Toru-母酵拟)、球(Saccharomyces rouxii母(Pe-菌球)、片(Weissella sp.菌氏斯)、魏cus sp.(Torulopsisversatilis母酵圆变)、易lopsis sp.)(Lactobacillus sp.菌酸)、乳diococcus sp.)(Zygosaccharomyces sp.母酵合)、结sp.母酵红)、胶(Candida sp.母酵丝假(Weissella菌氏斯)、魏(Lactobacillus sp.菌杆酸乳母酵赤毕)、(Rhodotorula mucilaginosa菌杆色)、无(Leuconostoc sp.菌珠串)、明sp.(Saccharomyces母酵酒)、酿(Pichia sp.)(Pediococcus sp.菌球)、片(Achromobacter sp.)cerevisiae(Gibberella sp.)菌霉赤母酵氏(Trichosporon asahii)、范母酵碱耐(Strepto-菌球(Lactococcus lactis)、链菌球乳酸乳母酵维鲁克斯克)、马(Vanderwaltozyma sp.球片酸)、乳(Lactobacillus sp.菌杆)、乳coccus sp.(Sac-母酵酒)、啤(Kluyveromyces marxianus(Bifi-菌杆歧)、双(Pediococcus acidilactici菌(Candida母酵丝)、假charomyces cerevisiae串)、明(Enterococcus sp.菌球)、肠dobacterium sp.)、(Saccharomyces unisporus母酵孢)、单sp.)(Leuconostoc sp.菌珠)(Pichia sp.母酵赤毕料原米、玉米、大米、糯豆、豌米、玉米、黍麦、小粱、高米、大粉乳或乳羊粱高麦小米稻米小米玉米小豆大菜蔬牛生酵发品食酒白酒黄醋食油酱菜泡乳酵发品制
1.2 酯合成霉菌的多样性
霉菌中关于红曲霉(Monascus sp.)产酯化酶催化小分子脂肪酸酯合成的研究最多[51],其次是青霉(Penicillium sp.)、根霉(Rhizopus sp.)、黑曲霉(Aspergillus niger)、米曲霉(Aspergillus oryzae)等。红曲霉能促进己酸、丁酸与乙醇的酯化过程,且产物以己酸乙酯为主,酯化能力极强,有文献报道以红曲霉生产酒用调香液,其己酸乙酯的含量稳定在8 g/L 的水平[52];根霉对己酸、丁酸和乙醇的酯化作用较弱,可催化生成己酸乙酯和丁酸乙酯[53]。
红曲霉嗜酸,耐高温,耐乙醇性较强,因此常应用于酿造和医药行业。杜礼泉[54]在红曲霉酯酶酯化特性的研究中发现该酶对乙酸、己酸、丁酸都有较强的酯化作用并生成相应的乙酯,其底物专一性较低,属于键合专一性的酶。红曲霉都可以以乳酸为底物,在浓香型白酒制作过程中,可以降低乳酸及乳酸乙酯的含量并催化己酸乙酯的合成,“增己降乳”利于白酒品质的提高[55]。烟色红曲霉能够分泌直接催化己酸和乙醇反应生成己酸乙酯的胞外酶,该酶适应的的温度范围较广,pH 值稳定范围大,使其在浓香型大曲酒生产中应用较多,能同时提高曲酒中四大乙酯的含量[54]。Guo 等[56]对来自红曲霉M-7 的冷活性酯酶Lip10 进行研究,发现该酯酶最适反应温度为40 ℃,对有机溶剂、金属离子有很好的耐受性,这些特性使Lip10 在食品工业和化学合成方面应用较为广泛。Kang等[57]从红曲霉M-7 中得到1 种高有机溶剂耐受性的酯酶Lip2,结果显示该酶作用的最适温度为40℃,且在60 ℃下孵育3 h,仍能保持70%的活性,对金属离子有较强的耐受性。在黄酒风味物质的合成过程中,红曲霉同样发挥重要作用。蔡琪琪[7]在研究红曲霉在红曲黄酒酿造过程中的作用时发现,酿造体系中红曲霉在发酵的前3 天是优势菌,并且红曲霉是有机酸的主要生产菌,可以促进酯的合成。
其它霉菌产酯化酶的研究亦有报道,而相比于红曲霉研究较少。张超等[8]对古越龙山发酵黄酒醪液中的微生物进行培养分离与鉴定,并使用GC-MS 对香味物质进行检测,发现青霉(Penicillium sp.)C01 产挥发性风味物质能力优于其它霉菌。我国酱油生产所用的菌种以米曲霉为主,米曲霉在酱油的发酵过程中能产生丰富的酶系,包括蛋白酶、淀粉酶、酯酶等,这些酶对酱油风味物质的合成具有重要作用[58];童佳等[59-60]、刘雅颀[61]研究发现以米曲霉作为主要菌种强化酱油发酵,酱醪中的特征性挥发风味物质包括己酸乙酯和辛酸乙酯等31 种酯类物质,且在发酵过程中,大量菌丝体发生自溶,菌丝体中含有丰富的蛋白质、核酸、糖类等,也对酱油风味有贡献。另外,通过大量的文献报道也发现根霉和黑曲霉等具有催化合成小分子脂肪酸酯的特性。例如:卢世珩等[62]从优质浓香型大曲中分离得到的根霉菌种Rh-9103,该菌生长快,繁殖力强,耐乙醇,温度适应范围大,主要产生催化合成己酸乙酯的酶类;孙舒扬[63]从白酒大曲中分离出的丝状真菌华根霉,其酯合成脂肪酶为SSL1 和SSL2,并发现SSL1 是在固态发酵中起重要作用的酯合成脂肪酶,该酶的最适温度为40 ℃,最适pH 值为8.0,纯化后对有机溶剂有较好的耐受性,对长链的脂肪酸酯催化活性较高。Guldhe 等[64]研究发现,黑曲霉来源脂肪酶具有高有机溶剂耐受性,可应用于小分子酯以及生物柴油的制备。
1.3 酯合成酵母菌的多样性
酵母菌是一类重要的酯合成真菌。从传统发酵食品的制作过程中分离出的酵母菌在增酯增香方面起到了重要作用。如应用于将白酒行业产生的大量丢糟中的有机酸转化成呈香酯类的南极假丝酵母(Candida antarctica)脂肪酶B,可提高酿酒副产物的利用率,节省成本,工艺条件为:南极假丝酵母脂肪酶B 用量0.2%(质量分数)、食用酒精(95%,体积分数)添加量20%(质量分数)、25 ℃反应8 d 条件下效果最好,用试验中所优化的工艺条件处理白酒丢糟能够提高丢糟中呈香物质的含量[65]。异常毕赤酵母中高产乙酸乙酯和乙酸的酵母菌株Ac-47 和Ac-19,分离于清香型白酒发酵过程中的酒醅,具有高效的乙酸乙酯催化合成特性[66]。刘飞等[67]从新疆哈萨克族奶酪中分离得到的4 株产脂肪酶的酵母菌菌株Y-2、Z-5、S-6、N-5,经鉴定Y-2、Z-5 属于马克斯克鲁维酵母(Kluyveromyces marxianus),S-6 属于毕赤酵母(Pichia sp.),N-5 属于乳酸克鲁维酵母(Kluyveromyces lactis),其中马克斯克鲁维酵母Z-5 的脂肪酶酶活可达25.99 U/mL。李学伟等[68]研究了添加生香酵母对酱油风味的影响,由于酵母发酵过程中产生酯化酶对有机酸类与醇类进行酯化作用,酯类物质在挥发性成分中的比例由0.99%提高到12.08%,显著提升了酱油的风味。龚加路等[18]从川南农家泡菜中筛选到1 株生香酵母Y7,其产生的主体挥发性香味是乙酸乙酯、丁酸乙酯等6 种物质,为泡菜发酵增香奠定了基础。
2 微生物源酯合成酶的多样性
传统发酵食品中存在酯合成能力的微生物种类十分丰富,对发酵食品的风味形成具有重要影响。微生物的酯合成特性,决定于微生物所携带的具有酯合成能力的功能酶。本节以乙酸乙酯、丁酸乙酯和己酸乙酯为代表性小分子脂肪酸酯,对微生物源酶进行概述。
2.1 合成乙酸乙酯的微生物源酶
乙酸乙酯的化学合成一般采用乙酸和无水乙醇在浓硫酸作用下生成乙酸乙酯,而化学来源的化合物在食品中应用的接受度越来越低,随着生活水平的提升,人们对天然来源的化合物的需求增大。通过微生物所产酶催化合成的化合物可被认为是天然化合物,因而需求逐步增大。微生物酶催化合成乙酸乙酯,具有反应效率高,条件温和易控,副产物少,操作简单安全,产品可被认为是天然化合物等优势。限制乙酸乙酯酶催化合成的主要因素包括底物的浓度、反应的最适条件以及酶的种类和用量等。葛清秀等[69]分离源自青霉FS1884-1 的碱性脂肪酶A,可在正庚烷中催化合成乙酸乙酯,当乙酸浓度为0.20 mol/L,乙醇浓度为0.25 mol/L,摇床转速为150 r/min,每间隔一定时间加入一定量的3A,分子筛移走产物中的水分,反应温度为36 ℃,反应时间为48 h,碱性脂肪酶A 的加入量为0.30 g/瓶 (相当于1 700 U/g 乙酸),此时催化效果最好,乙酸的转化率可达到71.4%。南极假丝酵母脂肪酶B 在其用量0.2%(质量分数)、食用酒精(95%,体积分数)添加量20%(质量分数)、25 ℃反应8 d 条件下酯化的效果最好,所得60%(体积分数)综合酒样品中乙酸乙酯质量浓度达到112.5 mg/100 mL[65]。Löser 等[70]研究了马克斯克鲁维酵母DSM 5422,该菌株所产酯酶可以催化合成乙酸乙酯。黄丹等[71]研究发现异常威克汉姆酵母(Wickerhamomyces anomalus)所产的酶可在低温条件下高产乙酸乙酯。郑树炀等[15]研究了山河陈醋生产中酵母菌产酯特性,分离得到的12 株菌株中,菌株QY18 发酵液中检出7 种特征性风味物质,其中乙酸乙酯含量为22.79%。对微生物法合成乙酸乙酯的研究越来越多,这奠定了该方法在我国传统发酵食品领域中的应用将更加广泛。
2.2 合成丁酸乙酯的微生物源酶
丁酸乙酯传统的化学合成方法是丁酸与无水乙醇在浓硫酸的作用下酯化,而反应体系中会含有少量的副产物醚等,应用于食品行业具有安全隐患,而利用微生物产酶合成丁酸乙酯则安全的多。产自于枯草芽孢杆菌的脂肪酶BSL2,可以在有机相中催化合成丁酸乙酯,以正己烷为溶剂,初始水活度为0.33,加酶量为100 mg,丁酸浓度为40 mmol/L,底物物质的量比(丁酸/乙醇)为1∶1.5,在40 ℃条件下振荡反应30 h,合成丁酸乙酯的转化率可达94.8%[15]。邢爽等[72]研究了浓香型、酱香型以及清香型大曲的酯化酶对合成丁酸乙酯的催化活力,研究发现:清香型和浓香型大曲的酶活力随丁酸浓度的升高先升高后降低,酱香型大曲的酶活力则是逐渐升高,清香型、浓香型和酱香型大曲的酶催化合成丁酸乙酯的最适pH 值分别为4.5,4.0 和3.0。不同微生物所产酶的催化特性各有不同,且需要考虑丁酸乙酯合成时其合适的酸浓度和pH 值范围。
2.3 合成己酸乙酯的微生物源酶
己酸乙酯是白酒中主要的香味物质,近年来,因为调制白酒行业的发展,己酸乙酯的需求量也随之增加,己酸乙酯的合成方法也在不断改进,微生物酯化酶催化己酸乙酯的合成是其中一种具有良好发展前景的方法,霉菌是主要菌种。王晓丹等[73]从生产现场分离得到的红曲霉I518 可以产催化己酸乙酯合成的脂肪酶,该菌株较理想的产酶发酵条件为:麸皮∶豆粕=2∶3,接种量20%,培养基初始pH 值为7,培养温度为32 ℃,培养时间为5 d,该条件下所产的粗酶制剂所催化的香酯液中己酸乙酯量达0.38 mg/mL,含量明显增加。烟灰色红曲霉所产酯酶的最适催化温度为40 ℃,最适催化酒精度为12%(体积分数),最适催化pH 值为3.0,属于键专一性酶,该胞外酶可以直接催化己酸和乙醇合成己酸乙酯[54]。另一种烟灰色红曲霉菌种Q-306,从实验室筛选的大曲中分离所得,该酯化红曲对己酸乙酯具有很强的催化专一性,在最佳催化温度为35 ℃,最适催化酒精度为20%vol,最适催化pH 值为3.5,最佳底物浓度为9.29 mg/mL条件下,酯化力为65 mg/(g·100 h)[74]。胡靖等[75]从贵州某知名酒厂浓香大曲中分离得到的紫色红曲霉IS518,该菌株的己酸乙酯酯化力较高,在实验室条件下可达62.79 mg/g,作为己酸乙酯脂肪酶产生菌有着较大的应用潜力。唐玉明等[76]对红曲酯化菌21-3 的特性研究中发现,四大酸和乙醇反应生成相应的乙酯,而四大酸的混合物与乙醇反应生成的物质却为己酸乙酯。己酸乙酯在传统发酵食品风味的形成方面,与其它小分子脂肪酸酯相比较为重要,因此天然健康的合成方法更加受到人们的重视。
此外,还有多种微生物源酶可以催化合成己酸乙酯。例如,郭通航等[34]从变质的食用油中分离得到1 株具有产己酸乙酯酯化酶能力的伯克霍尔德氏菌X10,研究发现该菌的最佳酯化条件为:己酸添加量2%,酯化温度35 ℃,酯化液初始pH 4.6,连续振荡48 h,优化条件下酯化酶活力由1.79 U/mL 提高到3.53 U/mL,较优化前增加了97.21%。在众多脂肪酶中,南极假丝酵母脂肪酶B 的用途最为广泛,它对非水溶性及水溶性物质都有很强的催化活性,尤其在非水相催化中,该酶表现出极好的活性和操作稳定性,该研究中利用α-凝集素表面展示系统成功的将南极假丝酵母脂肪酶B 锚定在酿酒酵母细胞的表面,并以该工程菌全细胞合成己酸乙酯作为应用模型,具有反应速度快,产率高,操作稳定的特点,当己酸与乙醇的摩尔比为1∶1.25 时,己酸乙酯的生成率为98.0%[77]。刘雪等[78]研究发现华根霉F8 发酵时可以产酯化酶,该酯化酶在一定条件下可以催化己酸乙酯的生成,在25 ℃下酯化25 d 可测得己酸乙酯含量为2 034.5 mg/100 mL。然而,由于中国传统发酵食品的多元性以及微生物的多样性,加之传统发酵食品现代化研究程度低,发酵食品来源聚焦于微生物酶资源序列和结构解析,以及催化机制的研究相对匮乏,也是食品领域今后的重要研究方向之一。
3 微生物酯合成酶的应用
3.1 发酵食品工业
发酵食品领域很多品类产品的风味与酯类物质密切相关。酯酶所具有的酯合成、酯水解[79-80]、酯交换[79]等功能,使其在发酵食品酯类合成中居重要地位(图1)。以白酒为例,中国白酒为世界著名的六大蒸馏酒之一,与其它蒸馏酒相比,中国传统发酵白酒拥有独特的风味,具有酯高、酸高、醛酮高、高级醇低的特点[81]。酯类中的己酸乙酯和乙酸乙酯等的含量和比例关系在很大程度上决定白酒的感官和风味[82]。以白酒中消费量最大的浓香型白酒为例,其感官及酒的品级,主要取决于己酸乙酯的含量以及与其它酯类物质的比例是否协调。酶制剂应用于白酒生产中,可以起到增加白酒酯类含量的作用,从而提高白酒的优质率。对于可应用到白酒领域的酯酶,首先要耐酸,其次对乙醇有良好的耐受性,并且酯化力强,以缩短酿造周期,提高生产效率。酯酶在其它传统发酵食品中也有应用,如酿造食醋、酱油、泡菜以及营养丰富的发酵乳制品。近年来,随着宏基因组文库、基因工程、定向进化技术、固定化技术的发展,以及界面酶促过程等技术的深入开展,高效产酯酶菌株的筛选、诱变育种、基因克隆及其定向进化构建高效基因工程菌的研究取得了良好成果,微生物酯酶的研究目前已经深入到分子层面,包括酶对底物的选择和作用机制[82],相关科学机制的解析,必将进一步有力推动酯酶在发酵食品工业的应用。
3.2 药物合成领域
化学合成法是医药领域很多药物合成的主要途径,然而许多药物都要求是手性化合物,这对化学合成是一个挑战。酶具有手性选择性,以酶作为工具拆分手性化合物已成为医药领域的研究热点,酯酶和脂肪酶是酶拆分中研究较为广泛的酶类[83]。Wen 等[84]考察了LipaseCAN、LipaseRH 2 种酶对α-苯乙胺的拆分效果,并与几种商品酶做了对照,结果表明,Novozyme435 来源商品酶的拆分效果最好。Godinho 等[85]用酯酶高效的对1,2-O 异亚丙基甘油酯进行拆分,其中对映体过量值可达到72%~94%,该酯酶还对1-乙酸苯乙酯有高对映选择性,得到具有>99%ee 的手性产物(R)-1-苯基乙醇,酶法拆分有效提高了药物的有效值。唐良华[86]研究凝结芽孢杆菌(Bacillus coagulans)ZJU318 和扩展青霉(Penicillium expansum)TS414来源脂肪酶催化拆分1 种常用的非甾体类抗炎手性药物布洛芬时发现,扩展青霉TS414 脂肪酶纯酶具有更高的催化活性和布洛芬对映体选择性(图2)。与其它化学方法相比,微生物源酶催化的反应具有条件温和、操作简便、安全,同时不会造成环境污染等优点,为实现工业化生产打下基础。又如前体药物中的酯类前药,在已上市的前药中,大约一半是经酯酶催化作用而产生活性。酯类前药常用来掩蔽水溶性药物中的羧酸、磷酸等带电荷基团,从而增加原药的脂溶性,提高它们的被动膜渗透性[87]。根据药物的水溶性不同,可以设计成不同的酯类前体药物,以适应其在体内发挥作用。

图1 酯酶的功能和在发酵食品中的应用Fig.1 The function of esterase and its application in fermented foods
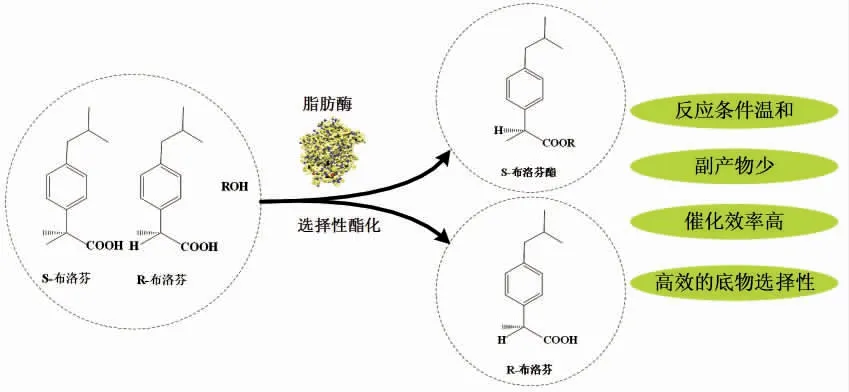
图2 酯酶在布洛芬【2-(-异丁基苯基)丙酸】对映体拆分中的应用Fig.2 Application of esterase in the resolution of Ibuprofen [2-(-isobutylphenyl) propionic acid]
3.3 化工领域
酶法合成小分子脂肪酸酯因其高效无污染,专一性强,杂质含量低,可适应连续化生产及产品分子质量易控制等优点[88],广泛应用于化工领域。如在精细化工中,可用酯直接作为表面活性剂,该做法可以提高效率,即增产,改进技术和质量;在精细化工中,酯类还可用作涂料中挥发性油漆的真溶剂,常用的有小分子脂肪酸酯类乙酸乙酯等;同时小分子脂肪酸酯还可应用于香料工业,例如:C8~C12酸与乙醇在酶催化作用下酯化生成的酯类,有玫瑰花香味,因此可用于玫瑰香精[89];还可应用于洗涤剂中,酯酶可以提高表面活性剂的释放,碱性脂肪酶可提高去污能力,而洗涤剂中酯类物质的存在可使产品呈现多种愉悦的香气[90]。
4 结论
近年来,许多研究从不同角度对发酵食品中微生物酯合成的过程进行解析,微生物合成的小分子脂肪酸酯类在食品、医药和日用化工等领域具有广泛的应用前景。酯酶和脂肪酶等羧酸酯水解酶类具有酯合成、内酯合成、酯交换等多种催化特性,酶催化不需要辅因子,并具有反应条件温和,方法简便,催化活性高,底物选择性强,产物易于分离等优点,且催化产物与化学合成方法相比可被认为是天然化合物,因而应用日益广泛。在传统发酵食品中,微生物(细菌、酵母菌、霉菌)合成的小分子脂肪酸酯是影响食品感官和风味的重要物质。微生物酯酶还是一种重要的工业酶类,虽然在产酶菌株的选育、培养条件、酶的性质及工业应用上已研究了几十年,但由于酶的结构及性质的多样性、酶的不稳定性、底物的水不溶性、酶的表达和提纯困难等问题[90-91],小分子脂肪酸酯的微生物合成仍有待研究和发展。随着微生物学、分子生物学、生物信息学和蛋白质工程技术的发展,新型微生物来源酯合成功能酶的种类必将不断丰富,结合更加有效和精准的酶定向改造技术,有助于获得底物专一性和键合专一性更强的酶,并推动其应用于特定种类的酯的合成和水解,以及化合物的精准高效手性拆分,产生重要的应用价值。
