黄淮与东北麦区小麦赤霉菌温度相关的致病力分化研究
2021-05-14雒丽丽张昊杨美欣王云飞许景升徐进姚强冯洁
雒丽丽 张昊 杨美欣 王云飞 许景升 徐进 姚强 冯洁
(1. 青海大学农林科学院 青海省农业有害生物综合治理重点实验室 农业农村部西宁作物有害生物科学观测实验站,西宁 810016;2. 植物病虫害生物学国家重点实验室 中国农业科学院植物保护研究所,北京 100193;3. 农业农村部国家植物保护甘谷观测实验站,天水741000)
由镰孢菌引起的赤霉病是危害麦类作物的一种重要病害,广泛分布于世界湿润和半湿润地区。小麦赤霉病是我国长江中下游麦区和东北春麦区主要流行性病害。近年来,在黄淮冬麦区也频繁流行成灾[1-3]。小麦赤霉病不仅造成产量损失,而且病菌产生的毒素污染小麦籽粒,危害人畜健康[1]。
温度、降雨和湿度等环境因素影响小麦赤霉病的发生流行,且在镰孢菌和其他真菌的生态学中起着重要作用[4-5]。在引起小麦赤霉病的多种镰孢菌中,禾谷镰孢菌(Fusarium graminarum)、黄色镰孢菌(F.culmorum)、燕麦镰孢菌(F. avenaceum)等为主要致病种[6],这些菌种分布广泛,其中禾谷镰孢菌在世界上较温暖、潮湿的地区(如北美和南欧)占主导地位[7],而黄色镰孢菌在较寒冷的地区如欧洲占主导地位[8]。小麦赤霉病原菌的种群组成与地理分布有关,病原菌因其地理生态环境差异在不同国家或不同区域的主要致病菌种类有所不同[8-9]。近年来,由于全球气候变暖、稻麦轮作以及免耕技术的推广,小麦赤霉病在全世界各产区呈扩张趋势[1]。不同区域病原菌组成也因气候变暖,品种更替及免耕技术而有所变化。
不同地理来源赤霉菌的致病力存在一定的差异。据报道,源自全国不同地区的禾谷镰孢菌菌株间均存在显著的致病力分化,且同一采样点采集的菌株间也存在明显的致病力分化[10]。Bai等[11]测定中国和美国的菌株在小麦上的致病力,来自中国的菌株的致病力明显强于美国菌株。研究发现禾谷镰孢菌的致病力差异与菌丝生长快慢有显著相关性[12],且禾谷镰孢菌的遗传分化及产毒类型对其致病力有一定的影响,Carter等[13]采用RAPD分析发现C组菌株对小麦和玉米的致病力显著强于A组和B组菌株,且A组产NIV毒素的菌株对玉米的致病力最强。因此明确不同地理来源赤霉菌群体的致病力分化对于病害的区域性防治策略的制定具有重要意义。
病原物对环境的适应性进化是其发生表型分化(包括致病力分化)的重要原因,其中温度的适应性是一个重要因素。据报道,导致我国松材线虫由低纬度向高纬度迅速扩张的原因除人为物流造成的长距离运输外,松材线虫本身对温度的适应性进化也是重要因素,其南北方种群已经形成了差异进化,北方种群耐寒性更强[14]。盛若成等[15]报道了低温条件下北方虫株 FCBX 对黑松的致病力强于南方虫株 AMA3。松材线虫种群在北移过程中经过了低温驯化,在低温条件下具有更强的繁殖能力[16]。Ellison等[17]利用“反向生态学”策略,对来自热带加勒比海和亚热带路易斯安那地区的粗糙链孢霉菌(Neurospora crassa)群体进行转录组测序,根据单核苷酸多态性位点系统发育分析,发现两地区的种群发生了明显的区域性分化。在全基因组范围内鉴定到两个群体“分歧岛”,其主要包含有温度和日照节律适应性基因,推测温度适应性是种群分化的重要因素。温度适合度实验证实,路易斯安那亚种群有更强的低温(10℃)适应性。
虽然在世界范围内对病原物温度适应性相关的致病力分化已有相关报道,但在镰孢菌该方面的研究信息较少,不同地理来源的镰孢菌群体的致病力与当地温度及适应性的关系尚不明确。鉴于此,本研究对采集自纬度差异较大的黄淮冬麦区和东北春麦区的小麦赤霉病病穗上的镰孢菌进行分离鉴定,明确病原菌的种群分布,研究不同温度条件下不同地理来源群体的致病力差异,为小麦赤霉病区域性防控策略的制定提供理论依据。
1 材料与方法
1.1 材料
2018 年和2019年间从黄淮冬麦区(河北省和河南省)和东北春麦区(内蒙古呼伦贝尔市)24个采样点采集小麦赤霉病穗。采用组织分离培养和单孢分离法[18]获得镰孢菌110株。所有菌株采用15% 甘油保存于-80℃ 超低温冰箱中备用。
1.2 方法
1.2.1 DNA的提取 供试菌株活化后,接种到YG培养基平板上(YG培养基:葡萄糖20 g,琼脂粉20 g,酵母粉5 g,蒸馏水1 000 mL,121℃ 高压灭菌 20 min),28℃ 黑暗培养5 d,收集菌丝于2 mL裂解管中,冷冻干燥后,低温研磨成粉末。基因组DNA的提取采用天漠生物公司的真菌/细菌RNA提取试剂盒。
1.2.2 镰孢菌种的鉴定 采用EF-1α基因序列比对和系统发育分析进行种的鉴定,扩增使用ef1和ef2引物[19]。PCR 反应条件 :95℃ 预变性 5 min ;95℃变性 30 s,53℃ 退火 30 s,72℃ 延伸 45 s,30 个循环;72℃ 延伸 10 min,10℃ 保存。PCR 反应体系 25 μL,其中 2× Taq PCR Mix 12.5 μL,12.5 nmoL ef1,12.5 nmoL ef2,DNA 模板62.5 ng,灭菌水10.5 μL。PCR扩增产物由生工生物工程上海有限公司进行单向测序,在Fusarium ID 数据库(http://isolate.fusariumdb.org/index.php)比对测序结果,一致性大于98%的确定是同一种。
优势单倍型检测:将优势群体F. graminearum菌 株 的 EF-1α序 列 利 用 MAFFT软 件[20]进 行序列比对,而后利用编写的Python脚本提取F.graminearum的单倍型,获得单倍型的种类及个数。
系统进化分析:从GenBank下载标准菌株参考序列,用MAFFT软件将本研究所有EF-1α单倍型序列与参考序列进行比对,而后采用ModelFinder计算最佳核苷酸替代模型(TIM2e+G4),iqtree软件最大似然法构建系统发育树,100次bootstrap[21]。系统发育树的编辑采用 treeio[22]、ggtree[23]和 Adobe Illustrator2020。
1.2.3 致病力测定 每个生态区各随机选取10株禾谷镰孢菌作为代表性菌株(共20株),在两个地点进行致病力测定。实验所用小麦品种为扬麦158。在中国农业科学院植物保护研究所廊坊实验基地以及青海大学西宁实验基地分别种植3个小区(2 m× 4 m)。廊坊为2019年10月播种,西宁为2020年3月播种。在小麦扬花初期进行接种实验,廊坊实验基地于2020年5月10日接种,西宁实验基地于2020年7月8日接种,每个菌株每小区接种5穗。日均温度由实验基地气象站记录。
分生孢子悬浮液制备:取少量菌丝于YG培养基中28℃恒温黑暗培养3 d,用6 mm打孔器打取菌饼接种于40% 绿豆汤培养基中[24],在180 r/min、28℃ 条件下摇培14 d,以血球计数板检测分生孢子浓度,8 000 r/min 离心后稀释或浓缩终浓度至1.0 ×106个/mL,保存于-80℃ 备用。
病原菌接种:采用单小花注射接种法。在小麦扬花期,选取长势相对一致的麦穗,在麦穗中下部小穗接种 20 μL混合均匀的分生孢子悬浮液,喷水雾后,套透明塑料袋保湿5 d。接种19 d 后调查病小穗率(病小穗率 = 每穗发病小穗数/每穗总小穗数)。两个生态区各随机选取10株禾谷镰孢菌作为代表性菌株,共20株菌,每个菌株在每个地点每小区接种5穗。
1.2.4 数据分析 采用t检验比较接种后21 d 两个实验基地日平均温度差异。采用广义线性模型比较黄淮和东北麦区赤霉菌群体致病力差异,数据分布型为二项分布。
2 结果
2.1 镰孢菌种的鉴定
2018 年和2019年间两个生态区共分离菌株110株,供试小麦赤霉病菌各采样点经纬度信息及分离菌株数量如表1所示。通过 EF-1α基因单倍型序列比对和系统发育分析,结果显示,所有菌株属于5个镰孢菌复合种,包括Fusarium graminearum species complex、Fusarium sporotrichioides species complex、Fusarium incarnatum-equiseti species complex、Fusarium tricinctum species complex 和Fusarium fujikuroi species complex( 图 1)。2018 年间两个生态区共分离出7个种共54株镰孢菌,F.graminarum为46株(85%),2019年间两个生态区共分离出6个种共56株镰孢菌,F. graminarum为50株(89%),表明F. graminarum是2018年和2019年间两个生态区的优势种。黄淮冬麦区分离出6个种共61株镰孢菌,F. graminarum为52 株(85%),F. asiaticum为4 株(6.7%),FIESC 为 2株(3.3%),F.verticillioides为1 株(1.6%),F. proliferatum为1株(1.6%),F. boothii为1株(1.6%)。东北春麦区分离出6个种共49株镰孢菌,F. graminarum为44株(90%),F. cerealis 为 1 株(2%),F. sporotrichioides为 1株(2%),F. avenaceum 为 1株(2%),F.culmorum为 1株(2%),FIESC为1株(2%)。 表明黄淮冬麦区和东北春麦区小麦赤霉病的优势种均为F. graminarum(表2)。
2.2 致病力的测定
2.2.1 接种后21d的日均温 接种后21 d,两地日均温差为-0.2-9.8℃,仅有一天西宁温度略高于廊坊0.2℃,日均温差平均值为4.7℃,t检验显示廊坊的日均温度显著高于西宁(P < 0.001)(图2)。
2.2.2 致病力测定结果 根据小麦病小穗率,不同菌株间致病力有明显的差异。廊坊接种结果显示,黄淮冬麦区和东北春麦区禾谷镰孢菌群体平均病小穗率分别为0.202和0.214(图3,表3),GLM分析表明两群体致病力无显著差异(P = 0.7)(表4),说明在较高日均温条件下,2个群体具有相似的致病力水平。西宁实验结果显示,黄淮和东北群体平均病小穗率为0.254和0.359(图3,表3),GLM检验表明两群体的致病力表现出明显的分化,在环境温度较低的条件下,东北麦区的禾谷镰孢菌群体的致病力水平显著高于黄淮麦区群体(P<0.001)(表4),表明东北春麦区禾谷镰孢菌群体在致病力方面具有明显的低温适应性。
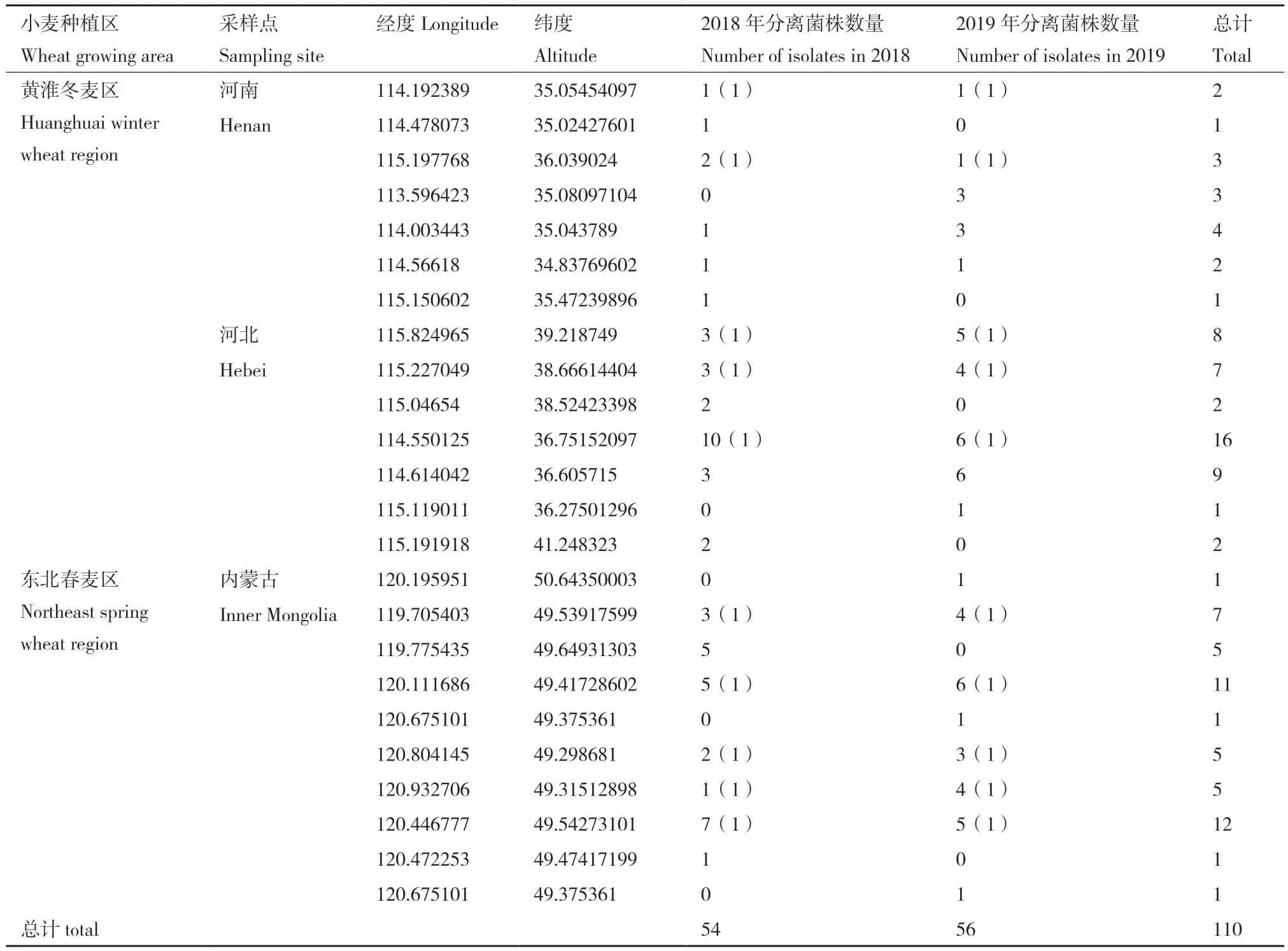
表1 供试小麦赤霉病菌信息Table 1 Information of WHB pathogens from wheat

图1 EF-1α 基因系统进化树Fig.1 Phylogenetic tree by EF-1α gene
3 讨论
本研究对2018-2019 年黄淮冬麦区和东北春麦区110株赤霉菌的群体组成进行了鉴定,共鉴定到5个复合种内共10个种的镰孢菌,两个生态区分别分离到6个种,说明这两个区域的赤霉菌组成较为复杂。其中F. graminarum(87%)是黄淮冬麦区和东北春麦区小麦赤霉病的优势种群。这与以前的研究结果一致[19,25]。F. sporotrichioides引起小麦赤霉病是在我国首次发现。两区域禾谷镰孢菌的致病力测定结果显示,廊坊实验站两区域禾谷镰孢菌群体致病力水平相似,而在西宁实验站,两个群体的致病力表现出明显分化,东北春麦区禾谷镰孢菌群体致病力明显高于黄淮冬麦区禾谷镰孢菌群体。
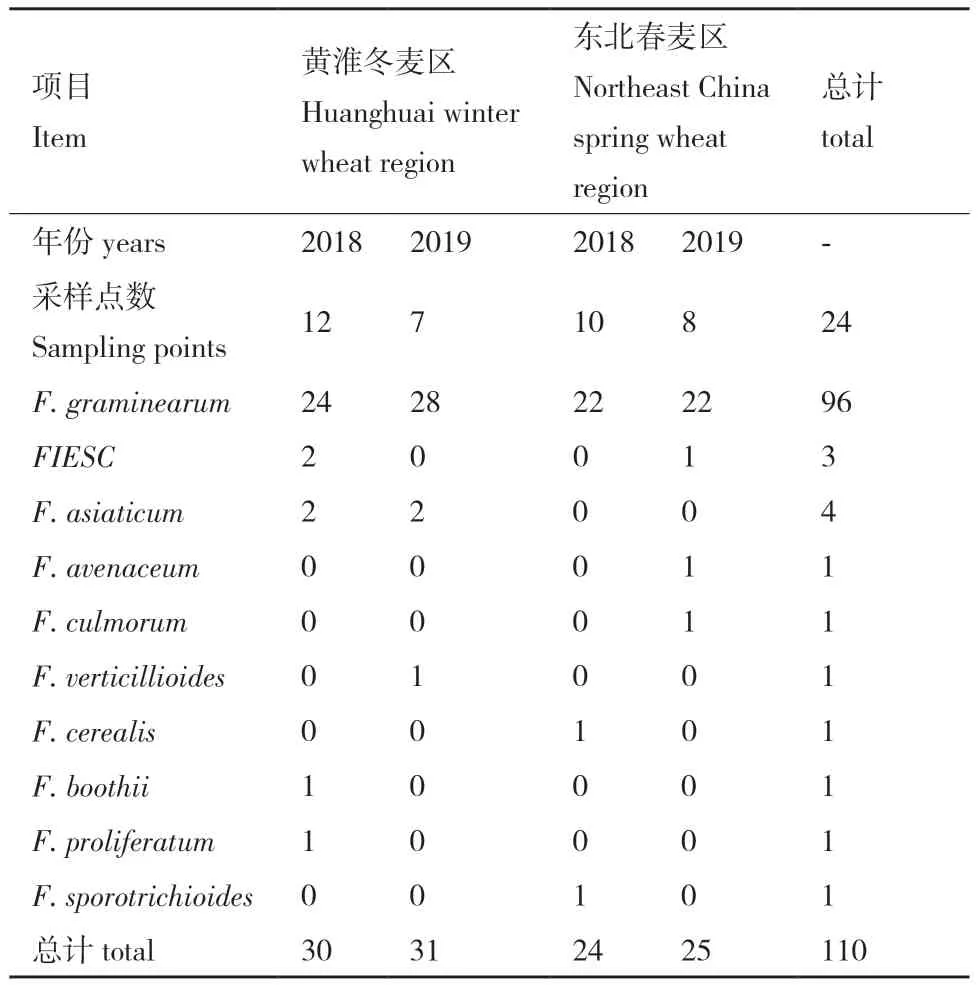
表2 2018和2019年间黄淮冬麦区和东北春麦区小麦镰刀菌种的分布数量Table 2 Number of Fusarium species on wheat from the Huanghuai winter wheat region and the Northeast spring China wheat region in 2018 and 2019
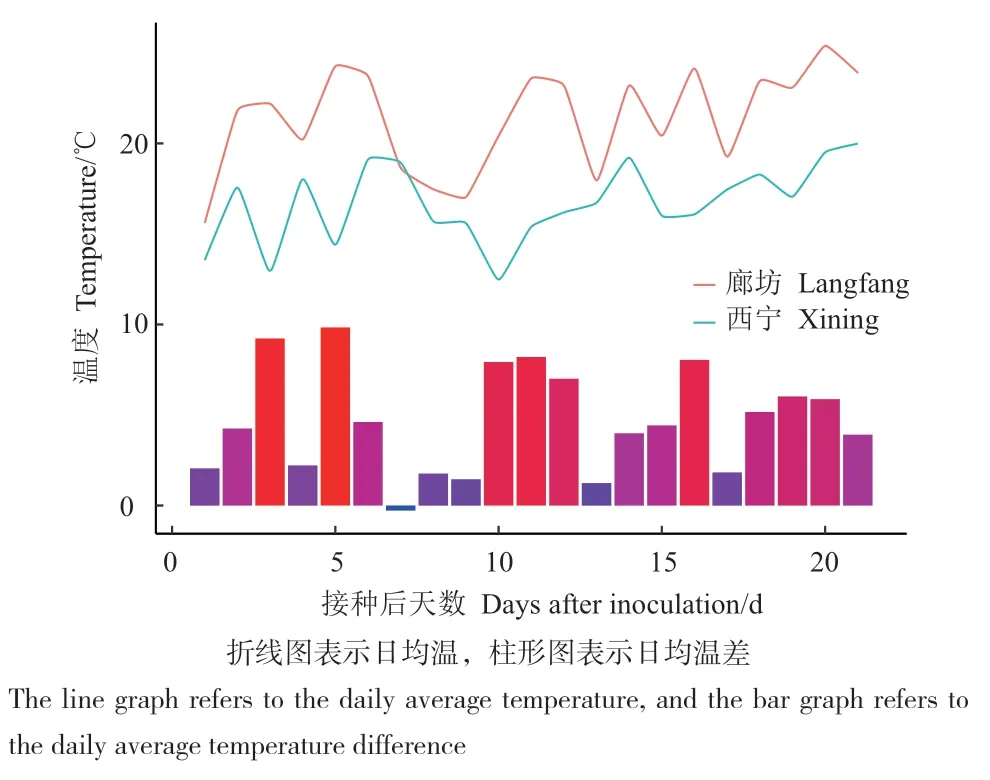
图2 接种后21 d日均温Fig. 2 Average daily temperature at 21 d after inoculation
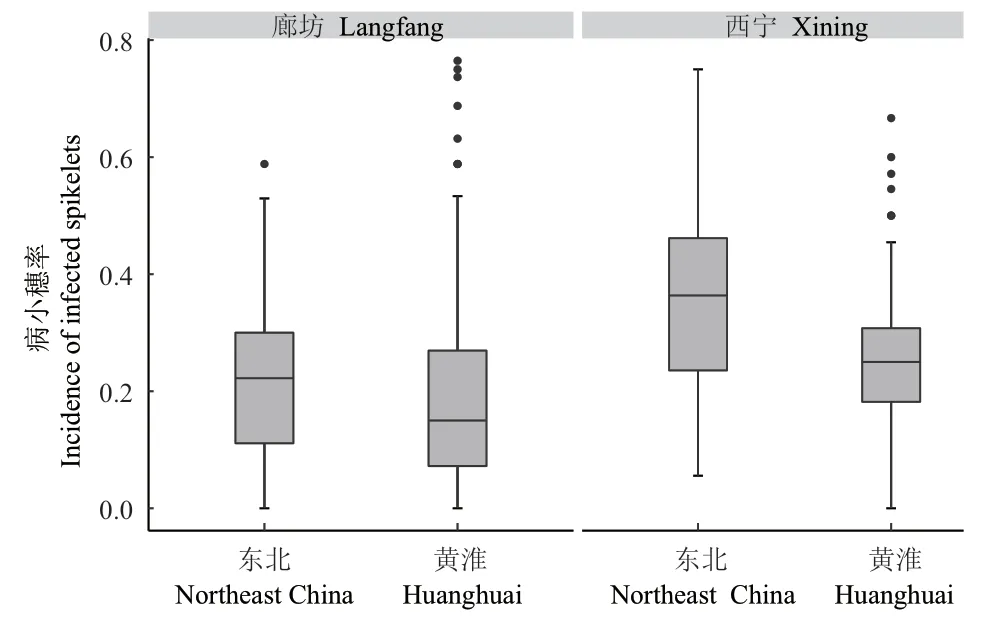
图3 致病力测定结果Fig. 3 Pathogenicity test results
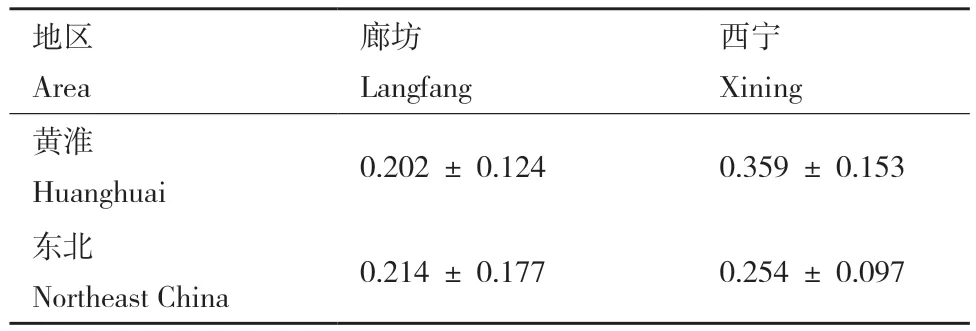
表3 黄淮和东北群体在两地平均病小穗率Table 3 Average incidence of diseased spikelets in Huanghuai and Northeast populations in the two sites
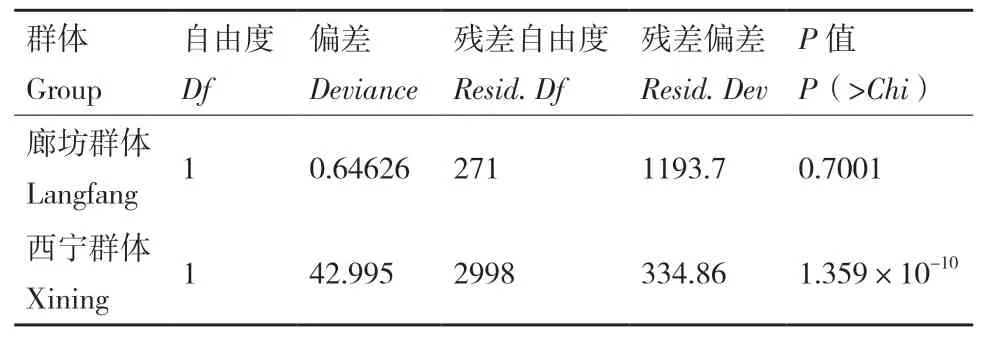
表4 广义线性模型比较群体间致病力差异Table 4 GLM analysis of pathogenicity between groups
致病力是植物病原菌侵染植物的重要表型,其强弱受到多种因素的影响。与寄主植物互作并发生协同进化是植物病原菌致病力发生变化的重要因素之一。如小麦条锈菌能够与当地主栽抗病小麦品种迅速发生协同进化,产生对抗病品种具有强毒性的新的生理小种,造成品种抗病性丧失。但目前尚没有小麦品种影响赤霉菌致病力方面的报道,中国抗赤霉病小麦品种主要种植在长江中下游麦区,而北方麦区主栽小麦均为感病品种[2],本团队前期研究显示,长江中下游麦区和北方麦区来源的小麦赤霉菌群体致病力没有显著差异[26]。本研究中黄淮和东北麦区虽然主栽品种不同,但均为感赤霉病品种,同时结合廊坊测定致病力无显著差异的结果,说明主栽品种可能并不是导致两区域赤霉菌群体在西宁表现出致病力差异的原因。杀菌剂选择压力也是影响病原菌致病力的重要因素,病原菌对杀菌剂敏感性的提高往往伴随着适合度的变化。Zhang等[27]发现对多菌灵产生抗性的禾谷镰孢菌群体具更高的适合度,其致病力和产毒能力更强。目前,长江中下游麦区小麦赤霉菌对多菌灵抗性问题严重,而北部麦区抗性频率很低[2]。其他常用杀菌剂如戊唑醇、氰烯菌酯等目前尚未检测到抗性菌株。同时,黄淮麦区和东北麦区赤霉病防治措施一致,均为为扬花期1次用药,因此杀菌剂压力可能也并不是造成两区域赤霉菌群体在西宁表现出致病力差异的原因。
镰孢菌对寄主植物的致病力受温度影响较大,不同镰孢菌种甚至同一种不同群体侵染寄主所需要的最适温度有所不同[28]。Maurin 等[29]报道雪霉叶枯菌(Microdochium nivale)侵染小麦需要较低的诱导温度(10-15℃),而禾谷镰孢菌和黄色镰孢菌侵染需要25℃以上的温度,因此,不同的镰孢菌种对温度的需求和适应能力存在差异,明确病原菌种的分布对研究病害的发生规律、调节温度等生态措施具有重要的指导意义。本研究中黄淮与东北麦区纬度相差较大,其环境温度具有较大差异,因此两区域的赤霉菌群体是否对温度具有一定的适应性是我们关注的重点问题。在环境温度较低的西宁实验站,东北春麦区禾谷镰孢菌群体致病力明显高于黄淮冬麦区禾谷镰孢菌群体,说明其具有明显的低温适应性。
不同地理来源的菌株往往具有与当地环境温度适应的特性,并进化出了相应的环境适应机制,因此在致病力上表现出不同的差异。Brennan等[30]报道了低温条件下雪霉叶枯菌对小麦幼苗的致病力较高,菌种和地理来源是影响其致病力的主要因素,雪霉叶枯菌分离株长期生活在冷凉多湿环境产生趋异适应,成为遗传上有差异的、适应冷凉环境的种群。Brennan等[30]还研究了温度对欧洲镰孢菌种的生长速率和致病力的影响,来自英国和爱尔兰的分离株通常在较低温度范围(10-25℃)下生长速率较快,而来自意大利和匈牙利的分离株在较高温度范围(20-30℃)下生长速率较快。这可能是由于镰孢菌分离株适应了各个来源国普遍存在的特定生长条件形成了温度生态类型。Wang等[31]研究了在不同温度(10℃、15℃、20℃和25℃)条件下与地下三叶草根腐病相关的真菌(F. avenaceum,Pythium irregulare,Rhizoctonia solani,F. oxysporum 和 Phoma medicaginis)的致病力,结果显示10℃条件下致病力最高,表明菌株地理来源也对其致病力有一定的影响,但程度不及菌种。Sabburg等[32]为进一步探究升高温度对镰孢菌致病力的影响,设置15/15℃,20/15℃,25/15℃和28/15℃四种昼夜温度处理,对11种小麦品种人工接种假禾谷镰孢菌进行致病力的测定。结果表明,小麦的茎基部褐变的程度以及病原菌定植的速度和DON的产生量均在15/15℃时最高,此研究验证了温度升高会降低病原体的适应性。上述研究均表明环境温度适应性对病原生物的致病力有重要影响。禾谷镰孢菌是小麦赤霉病最主要的病原菌之一,广泛分布于世界各地,已有较多报道显示禾谷镰孢菌菌株之间致病力差异明显,但尚未有致病力与地理来源显著相关的报道,这可能是以往研究选取的菌株菌处于相似的生态环境下。本研究选取黄淮冬麦区和东北春麦区地理距离南北相距约2 000 km,纬度相差超过15°,其环境温度具有显著差异,有利于本地病原菌发生适应性进化。研究发现两区域分离群体在低温下致病力分化明显,推测不同地理来源的菌株因遗传变异和自然选择的结果代表不同的基因型,所以即使将它们移植于同一生境,它们仍保持其稳定差异。东北春麦区禾谷镰孢菌种群由于对其本地气候条件有较强的适应性,从而在低温条件下致病力显著增高,其受特定天气条件的影响以及不断适应当地条件的自然选择作用下可能形成一套独特的与低温环境相适应的分子机制。
4 结论
本研究明确了我国黄淮冬麦区和东北春麦区小麦赤霉菌的种群组成,发现禾谷镰孢菌是这两个生态区的优势病原菌群体。同时,还发现不同温度条件、不同地理来源的禾谷镰孢菌群体致病力存在显著差异,证实了禾谷镰孢菌的致病力受地理分布、气候条件的影响,明确了赤霉菌的温度生态型。研究结果对于病害的区域性综合防治策略的制定具有重要意义。
