缺氮处理禾谷镰刀菌在氮素恢复后的致病力变化
2021-05-09柴喜存周天宇马海霞王世鹏刘玉涛
柴喜存,周天宇,马海霞,王世鹏,刘玉涛
(南京农业大学工学院,南京 210031)
0 引 言
秸秆还田具有明显改善土壤理化性质的作用[1-2],有研究表明其可以代替施用腐熟有机肥作为提升土壤肥力的有效措施[3];然而秸秆堆沤或过腹还田会增加收储等各环节作业成本1 095~1 380元/hm2,经济性较差[4];而直接原位还田又由于腐解缓慢、积温不足无法杀灭秸秆携入土中的病原菌,导致作物病害高发[5-7],这严重影响了农民秸秆还田的积极性,因而过剩秸秆仍是农业生产过程中亟待解决的问题[8]。
目前秸秆还田对植物病原菌的影响存在较大争议。Huang等[9]研究发现小麦秸秆、花生壳等7种有机物料还田后能在一定程度上抑制棉花枯萎病的传播;也有研究表明[10]通过有机物料的添加配合土壤pH值的调控及作物轮作,能够显著降低枯萎病的严重程度;但也有大量研究表明[11-12]秸秆还田是农作物病害高发的主要原因之一。研究认为有机物料的成分及腐熟程度[13]、微生物拮抗剂[14-15]、真菌毒性化合物[16-17]或营养[18]等的不同是作用效果出现较大差异的原因。目前关于氮素施用策略对病原菌产毒致病影响的研究较少,Cloud等[19]研究发现氮肥施用对菜豆壳球孢菌的定殖有显著性影响;Heier等[20]研究发现氮肥施用会提高赤霉病发生的严重程度以及真菌毒素的积累;但前述研究因缺少对关键指标的表征,存在因年份、气候等不同而结论差别较大的情况。秸秆还田会导致土壤内可利用氮素的量发生变化,Chai等[21]研究发现还田初期土壤内氮素缺乏能够提高病原菌的毒素产量和致病能力;而随着秸秆的分解和氮肥的补入,土壤中氮素含量提升对病原菌致病力的影响如何,尚缺少相关报道。
赤霉病(Fusariumhead blight,FHB)是严重影响小麦产量和品质的一种世界性病害,而随麦秸秆还田携入土中的禾谷镰刀菌是赤霉病的主要病原菌[22]。在中国长江中下游麦区,由于茬口紧,直接原位还田是小麦秸秆处置的重要方式,这为赤霉病的发生创造了条件[23]。因而本研究通过室内模拟小麦秸秆还田后土壤内有效态氮素的固持和释放过程,研究该过程对禾谷镰刀菌的毒素产量、致病能力、生长量及孢子萌发等的影响,以期阐明秸秆还田后病害高发的原因并为合理施肥策略的制定提供理论依据。
1 材料与方法
1.1 供试材料
禾谷镰刀菌菌株(Fusarium graminearum, F.graminearum)由南京农业大学植物保护学院周明国教授提供;小麦品种:鲁原502,赤霉病高度敏感,购自江苏省农业科学院;脱氧雪腐镰刀菌烯醇(Deoxynivalenol,DON)标准品为分析纯,购自SUPELCO(产品号:CRM46911,Bellefonte,PA,美国);其他化学试剂为分析纯,购自国药集团化学试剂有限公司。
基础培养基(MM-N):磷酸氢二钠12.8 g,磷酸二氢钾3.0 g,氯化钠0.5 g,氯化铵1.0 g,葡萄糖5.0 g,加入1 L去离子水,121 ℃高压灭菌20 min后加入2 mL 1 mol/L无菌硫酸镁和0.1 mL 1 mol/L无菌氯化钙溶液。
小麦培养基:将小麦粒于去离子水中浸泡2 h,再用去离子水煮30 min,风干5 h,最后使用粉碎机将小麦粒粉碎;取15 g小麦粒粉于100 mL锥形瓶中,115 ℃灭菌15 min。
1.2 试验设计及过程
本研究设置5个培养基N处理,调节MM-N培养基中氯化铵添加量以模拟环境中不同缺氮程度。5个处理氮素占比分别为0、25%、50%、75%和100%,其中100% 表示不缺氮处理,培养基中氯化铵浓度为1.0 g/L,其他处理按照比例添加相应量。每个N处理设置3个重复。
整个试验过程分为以下3个阶段:
1)菌丝培养:取0.5 cm×0.5 cm禾谷镰刀菌菌丝块接种至马铃薯葡萄糖琼脂液体培养基(Potato Dextrose Agar Liquid Medium,PDA)中培养72 h。
2)缺氮处理阶段:将菌丝经10 000 r/min离心10 min、无菌水冲洗后转移至5个不同N处理的MM-N培养基中培养168 h。
3)氮恢复阶段:收取缺氮处理阶段培养168 h后的菌丝,以10 000 r/min离心10 min,然后取相同量菌丝转移至MM-N全氮培养基中,并在0、2、24、48、72 h时取样,以用于生长量、毒素产量和侵染能力等指标的测定。
1.3 分析方法
1.3.1 禾谷镰刀菌生长情况测定
取5 mL菌液10 000 r/min离心10 min,去除上清液,使用天平称量湿菌丝质量,计算禾谷镰刀菌生长量。
式中M为单位体积(每毫升菌液中)菌丝的质量,g/mL;M1为菌丝和离心管的总质量,g;M2为离心管质量,g。
1.3.2 分生孢子萌发率测定
通过血球计数板对不同氮素水平处理168 h后的禾谷镰刀菌产孢数量进行计数;氮素恢复48 h后,取40μL菌液,在显微镜下对孢子萌发情况进行测定,分生孢子萌发率的计算公式如下:
式中Cgr为分生孢子萌发率,%;Cg为观察区域萌发的分生孢子个数;T是观察区域分生孢子的总数。
1.3.3 禾谷镰刀菌脱氧雪腐镰刀菌烯醇毒素产量测定
禾谷镰刀菌毒素的获取采用室内接种小麦粒的方式,利用高效液相色谱仪(HPLC,LC-20A,Shimadzu,日本)进行毒素测定[24];取0.2 g禾谷镰刀菌菌丝,加入3 mL无菌水后混匀,用于产毒及侵染能力的测定;取0.75 mL菌液接种至小麦培养基后,置于28℃恒温箱中培养10 d;然后进行毒素的提取和测定。
毒素的提取:小麦培养基中脱氧雪腐镰刀菌烯醇(Deoxynivalenol,DON)毒素的提取参考Femenias等[25]的方法并略有改进。取3.0 g小麦培养基,加入1.25 g聚乙二醇和25 mL去离子水,涡旋3 min,静置;再涡旋3 min,将 DON 完全浸出。以10 000 r/min离心10 min,取10 mL上清液以1滴/s的流速通过免疫亲和柱(产品号:IAC-030-3,PriboFast,中国);再用10 mL 磷酸盐缓冲溶液(Phosphate Buffer Saline,PBS)和10 mL去离子水先后淋洗免疫亲和柱,流速为1~2滴/s,弃去全部流出液,抽干免疫亲和柱;加入2 mL甲醇洗脱免疫亲和柱,收集全部洗脱液;在50℃下用氮气将洗脱液吹干,加入1.0 mL 18%(体积比)的甲醇水溶液,涡旋30 s以溶解残留物;经0.45μm微孔滤膜过滤,将滤液收集于进样瓶中备测。
毒素的测定:利用高效液相色谱进行测定,其工作条件:检测器为二极管阵列检测器(DAD,SPD-M20A,Shimadzu,日本);色谱柱为150 mm×4.6 mm的C18柱(产品号:5020-01731,GL Science,日本);流动相为18%(体积比)的甲醇水溶液;柱温为35 ℃;进样量为20μL;运行时间为15 min。
1.3.4 禾谷镰刀菌侵染能力测定
采取室内接种小麦苗的方式测定禾谷镰刀菌的侵染能力,以小麦苗的病变长度作为检测指标。取40μL上述菌液滴在脱脂棉上,并将吸有菌液的脱脂棉包裹在距离小麦苗(4~5 cm高)根部1.5 cm处的接种点,室温培养并保持脱脂棉湿度在65%左右;4 d后,使用数显游标卡尺测量接种位置的病变长度。
1.4 数据处理
研究中所有试验重复3次。利用Excel 2013、SPSS Statistics 25.0等软件进行数据处理分析;差异显著性检验使用Duncan法(α=0.05)。
2 结果与分析
2.1 缺氮处理禾谷镰刀菌氮素恢复后的生长和孢子萌发情况
生长及分生孢子的萌发在病原菌侵染作物的过程中具有重要的作用[26],禾谷镰刀菌缺氮培养168 h再氮素恢复后(0~72 h)菌丝生长情况如图1所示。由图可见,缺氮处理后的禾谷镰刀菌在氮素恢复2 h时,虽然各处理的单位体积菌丝质量(简称菌丝质量,下同)变化均较小,但已表现出明显差异,其中完全缺氮处理(即氮素占比为0)的菌丝质量为0.194 g/mL,显著高于其他处理(P<0.05);而氮素占比100%处理的菌丝质量最低(P<0.05),仅为0.127 g/mL菌液;在氮素恢复24 h时,各处理的生物量均有大幅度提升,完全缺氮处理的菌丝质量较转入初期提高了107.87%,达到了0.354 g/mL菌液,明显高于其他处理,说明完全缺氮处理在氮素恢复后具有很强的生长能力。随着培养的进行,由于过快生长对环境营养的消耗以及菌体死亡、自溶等原因,各处理间单位体积的生物量差别越来越小,培养至48 h时,各处理单位体积的菌丝质量已无明显差别;但在氮素恢复72 h时,氮素占比25%处理的单位体积的菌丝质量显著高于其他处理(P<0.05)。这一结果说明氮素恢复后由于生长速率等的不同会消除前期缺氮处理造成的生长差异。
不同氮素水平处理禾谷镰刀菌168 h后的分生孢子产量及氮素恢复后48 h孢子的萌发情况如表1所示。由表可见,完全缺氮处理的分生孢子产量最低,仅为8.2×105个/mL,显著小于其他处理(P<0.05);氮素占比75%处理的分生孢子产量最高,达到了4.18×106个/mL,显著高于其他处理(P<0.05)。不同处理间分生孢子的萌发率有明显差别,其中完全缺氮处理产生孢子的萌发率显著高于其他处理,为46.00%(P<0.05);而不缺氮处理产生的孢子萌发率最低,仅为3.00%(P<0.05)。由分生孢子的数量与萌发率相乘,得到5种不同处理萌发的孢子数量分别为377 200、698 754、159 536、320 606和87 900个/mL,这意味着如果不考虑原有菌丝的活力,则完全缺氮处理和氮素占比25%处理在氮素恢复后具有较大的生长潜力。这充分说明不同缺氮处理对氮素恢复后禾谷镰刀菌的孢子萌发及生物量增长具有重要影响。
从氮素恢复后的孢子萌发以及生长情况来看,不同程度缺氮处理在氮素恢复后具有明显不同的生长速率,高的分生孢子萌发率可能是前期完全缺氮处理生长量较大的原因。
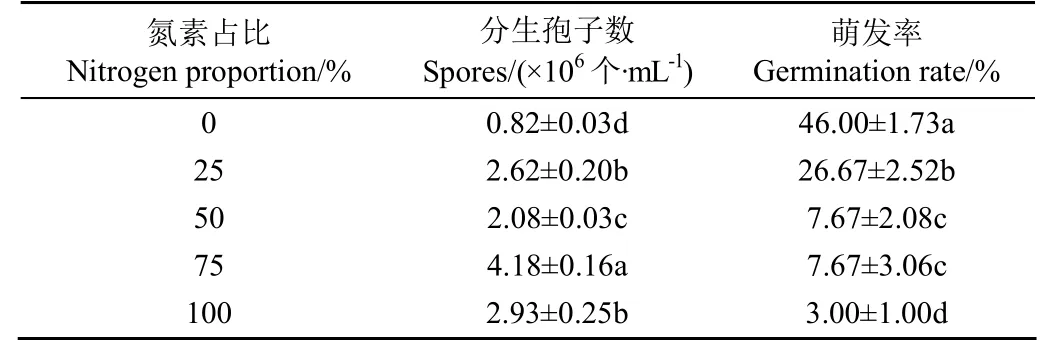
表1 缺氮处理对禾谷镰刀菌分生孢子产量及萌发率的影响Table 1 Effects of nitrogen deficiency treatments on production and germination rate of F. graminearum spore
2.2 缺氮处理禾谷镰刀菌氮素恢复后产毒能力的变化情况
DON是禾谷镰刀菌产生的真菌毒素之一,能够促进镰刀菌对禾本科植物的侵染[27],研究以DON的含量为检测指标,来确定氮素恢复对缺氮处理禾谷镰刀菌产毒能力的影响。氮素恢复后禾谷镰刀菌的产毒能力变化情况如表2所示,由表可以看出,完全缺氮处理(即氮素占比为0)在氮恢复初期的毒素产量较高,达到了0.200μg/g;恢复至2 h时,未检测到DON毒素;而后毒素的产量略有提升;但在整个氮素恢复的72 h内,完全缺氮处理的产毒能力较低。氮素占比25%处理在氮素恢复24 h时DON毒素产量达到1.361μg/g,显著高于同时间的其他处理(P<0.05);氮素占比50%、75%和100%处理均在氮素恢复后72 h毒素产量最大,分别达到了0.515、1.409和0.540μg/g。这一结果说明完全缺氮处理禾谷镰刀菌氮恢复后的产毒能力受到了明显的影响,而其他处理则在氮消耗到一定程度时达到最高产毒能力。

表2 不同缺氮处理氮素恢复后0~72 h禾谷镰刀菌的产毒量Table 2 Deoxynivalenol (DON) production of F. graminearum under different nitrogen deficiency treatments from 0-72 h after nitrogen recovery
2.3 缺氮处理禾谷镰刀菌氮素恢复后侵染能力的变化情况
侵染能力与病原菌的致病力密切相关,研究以不同程度缺氮处理的禾谷镰刀菌氮素恢复后侵染麦苗的长度作为判断侵染能力的依据,其变化情况如表3所示。由表可以看出,完全缺氮处理(即氮素占比为0)禾谷镰刀菌168 h后即氮恢复开始时,其侵染能力较弱,侵染长度仅为4.19 mm,显著低于其他处理(P<0.05);24 h后随新生菌丝的增多,其侵染能力得到一定程度的提升,至48 h时,侵染长度达到了8.93 mm;氮素恢复72 h时,小麦苗病变长度为7.73 mm,显著低于氮素占比25%、50%和100%处理(P<0.05)。氮素占比100%处理在氮恢复的初期侵染长度较高,达到了13.28 mm;而后在2~48 h的侵染长度大幅度下降,在72 h又上升为11.96 mm。氮素占比25%、50%和75%这3个缺氮处理均在氮恢复后的24 h时侵染长度达到最低,而后又提升,至72 h时达到最大值。这说明不同氮素占比处理的禾谷镰刀菌在氮素恢复的初期侵染能力会有所降低;而随着氮素的消耗,其侵染能力会提高。
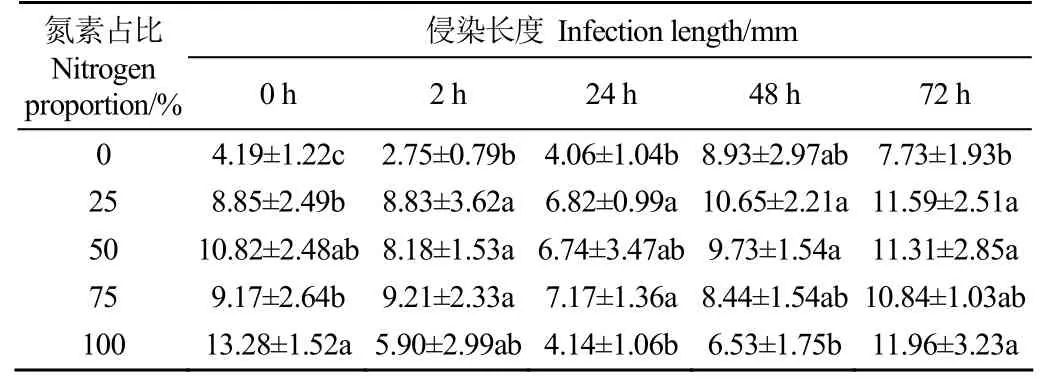
表3 不同缺氮处理氮素恢复后0~72 h禾谷镰刀菌的侵染长度Table 3 Lengths of infected lesions caused by F. graminearum under different treatments from 0-72 h after nitrogen recovery
3 讨 论
大量研究表明氮元素和植物病害及其抗性间有着密切的关系[28];秸秆还田能够改变土壤碳氮组成和土壤微域环境,从而影响其中微生物的种群结构及生理功能[29]。由于微生物分解秸秆需要从土壤中吸收一定量的氮素[18],因此单纯秸秆还田会提高土壤氮素的矿化速度[30],显著降低土壤铵态氮和硝态氮的含量[31],降低耕层土壤氮的有效性[32]。有研究表明不同形态的氮素能够影响香蕉枯萎病的发病率和严重程度[33],但不同报道间结果并不完全一致[28]。本试验着重研究了氮素缺乏对禾谷镰刀菌致病性的影响,发现在氮素完全恢复后,经受完全缺氮处理168 h的病原菌其分生孢子萌发率达到46.00%,显著高于其他氮素水平,这可能是由于长时间的缺氮处理耗尽了病原菌内的含氮化合物,使得外源氮素成为了孢子萌发的必要条件导致的[34]。研究对秸秆还田后土壤内氮素的供应情况进行了细分,发现完全缺氮处理病原菌的产毒量以及侵染小麦苗的长度处于较低水平;部分缺氮或不缺氮处理在氮素恢复的初期,产毒和致病能力均有所降低,但随着培养的进行,由于培养基中氮素的消耗,会使得禾谷镰刀菌产毒和侵染能力再度得到提升;这能很好解释Heier等[20]发现的氮肥施入对赤霉病作用效果迥异的原因。研究结果充分说明秸秆还田后氮素含量变化在病原菌致病力变化中起着非常重要的作用,因而从生产实际来看,秸秆还田初期施入充足的氮肥或后期追施足够的氮肥,将有利于病害的控制。
以往研究对氮素与病原菌致病力间的关系得到了不同甚至完全相反的结论,有学者提出可能是氮素形态、病原菌类型以及气候等因素的不同导致了上述差异[28],却忽视了氮素变化对病原菌自身生理的影响。秸秆还田后的氮素缺乏环境极易诱导病原真菌发生细胞自噬[35],而细胞自噬是表征真核生物响应微域环境可利用氮素的重要指标,有研究也发现秸秆腐解会诱导里氏木霉的7种自噬相关基因表达上调[36]。因而,与氮素代谢密切相关的细胞自噬在禾谷镰刀菌致病力变化中的作用有必要进一步阐明,这有可能为秸秆还田后植物病害的控制提供崭新思路。
4 结 论
1)氮素恢复后,完全缺氮处理和氮素占比25%处理的分生孢子具有较高的萌发能力,菌液中萌发的孢子数量分别为377 200、698 754个/mL,高于其余处理;高的孢子萌发率是完全缺氮和氮素占比25%处理生长量较大的原因。原有菌丝的活力能够保持部分缺氮处理(即氮素占比为50%和75%)和不缺氮处理的生长潜力;因而氮素恢复能够消除不同程度缺氮处理对禾谷镰刀菌生长的影响。
2)禾谷镰刀菌在氮素恢复后,其产毒和侵染能力会短暂下降;但随着氮素的消耗,禾谷镰刀菌的产毒和侵染能力会再度得到提升;表明恢复后氮素的消耗情况对禾谷镰刀菌的侵染能力具有较大的影响。
本研究涉及的缺氮处理与氮素恢复处理,分别与还田后秸秆腐解驱动的土壤内氮素固持与释放相对应;研究发现的氮素恢复后经缺氮处理禾谷镰刀菌的产毒和致病力变化规律,为秸秆还田后合理肥筹策略的制定提供理论依据:即在秸秆还田过程中施入或追施充足的氮肥,缩短病原菌处于部分缺氮状态的时间。
