可可毛色二孢效应子LT_397基因的克隆及转录分析
2021-04-30曹阳邢启凯李铃仙徐品三
曹阳 邢启凯 李铃仙 徐品三


关键词可可毛色二孢;LT-397基因;生物信息学;逆境胁迫;过敏反应
葡萄属于葡萄科葡萄属木质藤本植物。葡萄因具有营养价值高、易加工等特点,深受人们的喜爱。2018年,我国葡萄的种植面积约87万hme(国家葡萄产业技术体系)。但葡萄病害长期制约葡萄产业的健康发展,其中由葡萄座腔菌科Botryosphaeriaceae真菌引起的葡萄溃疡病(Bot-ryosphaeria dieback)是一种重要的葡萄枝干病害。该病害引起枝干溃疡、果梗干枯、掉粒等病征,严重时会造成整株植株枯死。在我国葡萄产区共发现6种葡萄座腔菌科真菌可引起葡萄溃疡病,分别是Botryosphaeria dothidea、Diplodia seriata、Lasiodiplodia theobromae、Neofusicoccum par-rum、L.pseudotheobromae和Neofusicoccumman-gi ferae,优势病原菌为葡萄座腔菌B.dothidea和可可毛色二孢L.theobromae,致病性最强的是可可毛色二孢。目前对葡萄溃疡病的研究主要集中在病原真菌的分离鉴定、种群结构和化学防治等方面,对主要致病菌可可毛色二孢L.theobromae的致病机制尚不明晰。
在植物与病原物的长期协同互作过程中,植物形成了多种免疫机制来阻止病原物的侵染;同时病原物也进化出多种致病机制来克服宿主的防卫反应,最终使宿主发病。效应子是一类由病原菌产生,通常富含半胱氨酸的蛋白分子,可分泌到寄主植物细胞问或细胞内,抑制寄主的防御反应,从而促使病原菌在寄主植物上定殖、侵入和扩展。已发现真菌效应子主要通过靶标蛋白来影响植物的PTI(PAMP triggered immunity)防御反应,如小麦条锈菌Puccinia striifrmis f.sp.tritici效应基因PEC6可以与拟南芥和小麦中的腺苷激酶互作,从而抑制寄主PTI,促进小麦条锈菌在寄主中的侵染。稻瘟菌Magnaporthe oryzae效应基因SLP1与番茄叶霉病菌Cladosporiumfulvum效应基因ECP6含有保守的LysM结构域,能与植物几丁质结合受体竞争结合几丁质,从而逃避PTI防御反应。植物激素对调控植物的防御机制起着重要的作用,真菌效应基因也通过影响植物激素的合成来调控植物的防御反应。例如玉米黑粉菌Usti-lago maydis分泌的效应子Cmul能够催化水杨酸的前体物质分支酸转为预苯酸(prephenic acid),抑制水杨酸的生物合成,影响水杨酸信号传导途径。某些真菌效应子通过干扰宿主植物的活性氧生成来抑制植物的防御反应,玉米黑粉菌的效应子Pepl直接与玉米的过氧化物酶POX12互作抑制活性氧介导的免疫反应。病原真菌效应蛋白也可以通过抑制RNA沉默来抑制植物的防御反应,小麦秆锈菌P.graminis f.sp.tritici效应子PgtSRl通过改变防御调节因子小RNA的丰富度来抑制植物RNA沉默诱导的防御机制。总之效应子可以通过不同的信号途径来抑制寄主的免疫反应,从而使病原物能够完成侵染、定殖寄主植物的过程。Yan等对L.theobromae进行全基因测序组装分析,并结合生物信息学方法预测发现:共359个具有效应子特征的基因。但目前对可可毛色二孢效应子的功能研究还未见报道,效应子与寄主葡萄之问的互作机制并不明晰。
探究效应子基因的转录模式及致病性对研究病原菌效应子与寄主植物的作用机制有重要意义。本试验对可可毛色二孢特有的效应子基因LT_397进行克隆,得到目的基因ORF全长并进行生物信息学分析,对LT_397基因受可可毛色二孢诱导转录分析和逆境胁迫转录分析,利用烟草毒力测定体系对效应子LT_397进行毒力测定。研究结果为进一步分析病原菌对寄主的作用机制提供了理论基础。
1材料与方法
1.1试验材料
可可毛色二孢菌株CSS-O1s由本实验室分离并保存。水稻谷枯病菌(颖壳伯克氏菌)Burkholderiaglumae菌株106619和pEDV5载体由中国农业大学孙文献课题组馈赠。本氏烟Nicotianabenthamiana培养条件为26℃,L//D=14 h//10h,取生长4周烟草进行接种试验。
完全培养基(cM):6g/L酵母提取物、3g/L酪蛋白、10g/L葡萄糖和0.1%(V/V)微量元素混合液;基礎培养基(MM):6g/L NaN03、0.5g/LKC1、1.5g/L KH2P04、0.5g/L MgS04、10g/L葡萄糖;缺碳源基础培养基(MM-C):6 g/L NaNO3、0.5g/L KCl、1.5g/L KH2P04、0.5g/L MgS04;缺氮源基础培养基(MM-N):0.5g/L KC1、1.5g/LKH2P04、0.5g/L MgS04、10g/L葡萄糖;培养基pH均调至6.5。
LA Taq DNA聚合酶、pMDl8-T载体、SYBRPremix Ex TaqTM购自宝生物工程有限公司。Trizol和反转录酶SuperScript*Ⅲ购自Invitrogen公司。大肠杆菌Escherichia coli Trans5a感受态购自全式金生物技术有限公司。其他常规化学药品均为分析纯。
1.2试验方法
1.2.1 LT 397基因的克隆
采用TRIzol法(Invitrogen)提取可可毛色二孢用DNA凝胶回收试剂盒对目的基因进行纯化回收,并与pMDl8-T载体连接,转化大肠杆菌Trans5a,挑取单克隆进行菌液PCR验证,阳性克隆送至北京擎科新业生物技术有限公司进行测序。
1.2.2效应子LT397的生物信息学分析
利用Edit Seq和ExPAsy软件对LT_397进行氨基酸序列分析;Signal P 4.1 Server预测LT_397氨基酸序列信号肽;使用Protscale和SOPMA软件测定LT_397跨膜螺旋等二级结构和蛋白质表面氨基酸分布;Swiss-Model预测LT_397蛋白的结构模型。
1.2.3逆境胁迫处理对LT_397基因转录水平的影响
检测不同胁迫处理条件下可可毛色二孢CSS-01s效应子LT_397基因的转录情况。营养胁迫处理:将CSS-01s菌株于CM液体培养基上26℃160 r/min摇培24h,离心收集菌丝,随后将菌丝分别置于缺碳源MM-C和缺氮源MM-N培养基上生长4h。氧化胁迫处理:将CSS-01s在CM液体培养基上培养24h后,向培养基里分别加入HzOz至终浓度为5mmol/L和10mmol/L处理4h。高温胁迫处理:CSS-01s在CM液体培养基上培养24h,转移到42℃培养箱静止45min。离子胁迫处理:CSS-01s在CM液体培养基上培养24h,离心收集菌丝,分别转移到浓度为1mol/L NaCl和浓度为0.1mol/L LiCl的MM培养基中处理4h。酸碱胁迫处理:CSS-01s在CM液体培养基上培养24h,离心收集菌丝,转移到pH分别为4和8的MM培养基中处理4h。紫外线UV胁迫处理:将CSS-01s在CM液体培养基上培养24h后,用UV-B辐射测定仪(北京师范大学光电仪器厂)进行测定,辐射强度为5kJ/m2,分别设处理时问为5min和10min。处理完毕后,将离心收集的菌丝置于液氮中速冻,-80℃保存。
1.2.4qRT-PCR检测LT_397基因的转录水平
液氮研磨菌丝后,采用TRIzol(Invitrogen)法对处理样本RNA进行提取。RNA经DNase I(TaKaRa)消化处理后,运用SuperScriptⅢ(Invitrogen)反转录合成第一链cDNA为模板进行实时定量PCR扩增(qRT-PCR)。设计qRT-PCR特异性引物,LT_397基因(登录号GenBank:KAB2577062.1)(F:5-CAT-CACCTACACCACAC-3;R:5'-CTCCTTGTCG-TATTCAGTCT-3);LtActin基因(登录号GenBank:AY846879.1),(F:5'-TCTTCGCFCGAGAAGTCC-TA-3;R:5-ACAATCG-AACGTCCTC-3)。参照TaKaRa说明书,采用SYBR Green I染料法进行荧光定量PCR,反应在ABl7500 Reabtime PCR检测系统上进行。PCR反应体系:10μL SYBRPremix Ex TaqTM、0.5μL上游引物(10μmol/L)、0.5μL下游引物(10μtnol/L)、0.4μL ROX Refer-enceDye II,0.5μL cDNA模板、补水至20μL。反应程序为:95℃预变性3 min;95℃30 s,60℃30 s,72℃30 s共40个循环。每批检测3次,设3个生物学重复。根据实时荧光定量PCR得到的ct值以及标准曲线,采用2法分析试验数据。以接种可可毛色二孢时问0h作为参照(转录水平为1),计算LT_397基因受可可毛色二孢不同侵染时间的相对转录水平。以未做胁迫处理的样本CK作为参照(转录水平为1),计算LT_397基因逆境胁迫处理时的相对转录水平。
1.2.5效应蛋白LT_397毒力测定
以397-T为模板,用引物397F(5-ACTC-GAGGGCCCTGTGGAAAAGCGTC-3)和397R(5'-AGGATCCGTTGGCGTCTTCGTAAACG-3)进行PCR扩增(扩增反应1.2.1),扩增得到的片段连接到T载体上,得到的重组载体命名为397XB-T。由北京擎科新业生物技术有限公司进行测序验证。用Xho I和BamHI双酶切397XB-T载体,回收小片段插入到pEDV5载体的Xho I和BamHI酶切位点间,得到的重组载体命名为pEDV-397。
参照Sharma等和Zhang等的方案,挑取颖壳伯克氏菌单菌落接种于LB液体培养基,28℃振荡培养至0D=0.8。4℃下800g离心10min,弃去上清。用25mL预冷的10%甘油溶液轻轻悬浮细胞,4℃下800g离心10min,弃上清,重复上述步骤一次。最后加入700μL预冷的10%甘油溶液悬浮细胞,即为颖壳伯克氏菌感受态细胞。采用电激转化方法,将pEDV-397载体转入颖壳伯克氏菌感受态细胞。取200μL培养液,涂布于含25mg/L庆大霉素(gentamicin)的LB固體平板上,28℃倒置培养筛选获得转化子,并利用PCR验证转化子是否构建成功。
将转化子和空载菌株接种于10mL含有25mgLGM的LB液体培养基中,28℃,180r/min振荡培养过夜。用0.9%NaCl调整OD=0.1。用1mL注射器将菌液注射人烟草叶片下表皮。2~3d后观察细胞坏死情况。
2结果与分析
2.1 LT_397基因PCR结果分析
设计特异性引物,以可可毛色二孢的cDNA为模板,克隆LT_397基因,测序分析结果表明,LT_397开放阅读框(ORF)全长1140 bp(图1)。
2.2 LT_397蛋白的基本性质与功能分析
2.2.1 LT_397蛋白的氨基酸序列分析
以NCBI中基因登录号KAB2577062.1序列为参照序列,利用ExPAsy在线软件分析LT_397的开放阅读框。如图2所示:LT_397基因是编码379个氨基酸的多肽。Edit Seq软件分析该多肽相对分子质量约为40.8 kD,有17个碱性氨基酸、51个酸性氨基酸、131个疏水性氨基酸和137个极性氨基酸,含有11个Cys。等电点为3.79,在pH为7时,电荷为-34.06。
2.2.2 LT_397信号肽分析
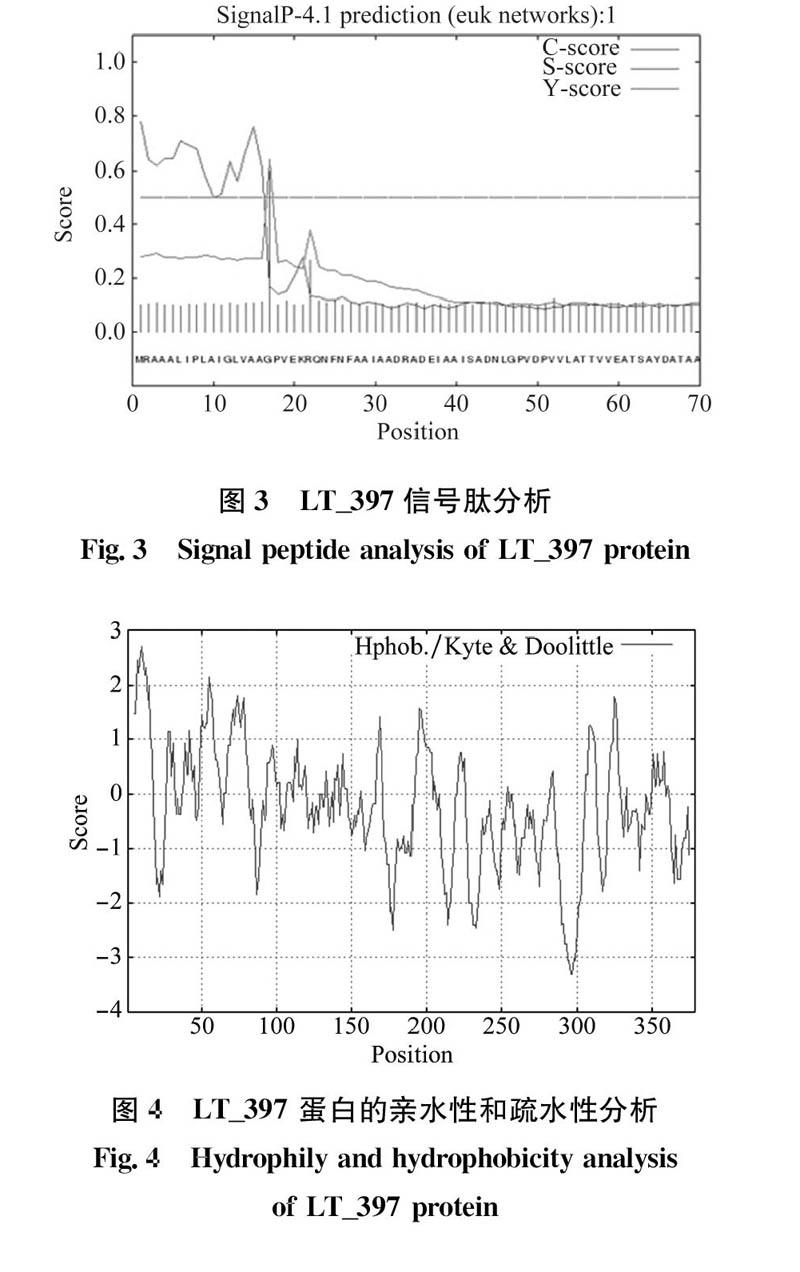
信号肽指的是一段通常位于蛋白质N端,在新合成的多肽链中指导蛋白质合成及跨膜转移,大小为15~30个的氨基酸多肽。它一般由3部分组成,即疏水核心区、信号肽的c端和N端。使用Sig-nal P 4.1 Server在线软件对可可毛色二孢效应子LT_397蛋白序列进行分析,预测其信号肽由16个氨基酸组成(图3)。
2.2.3 LT_397蛋白的亲水性和疏水性分析
维持蛋白质结构特征的主要因素是疏水和亲水问的平衡。疏水作用有利于与氨基酸的基团形成氢键等作用力使蛋白质结构稳定,因而蛋白质的稳定性很大程度上依赖于分子内的疏水相互作用。使用在线软件Protscale分析LT_397蛋白分子的亲水性和疏水性,并根据蛋白的得分数(Score)来计算蛋白的亲、疏水性。結果表明,该蛋白的氨基酸Score值在3~-3.5(图4)。疏水性质用正值来表示,亲水性质用负值表示,且绝对值越大,亲、疏水的性质则越明显。用Protscale工具统计分析显示GRAVY值为-0.244(最小值:-3.300,最大值:2.711),表明LT_397为具有较强亲水性的蛋白质。LT_397蛋白前15~20个氨基酸均为疏水性氨基酸,表明在N端有一典型的疏水区域,可能与信号肽的组成相关。
2.2.4 LT_397蛋白二级结构分析
使用在线软件SOPMA对LT_397蛋白的二级结构进行分析,结果显示LT_397氨基酸序列的二级结构是由31.13%的a螺旋、15.83%的延伸链区和49.39%的无规则卷曲并通过3.43%的β转角进行连接构成。
2.2.5 LT_397蛋白三级结构分析
使用Blast P对LT_397蛋白进行同源序列比对,选择同源序列E值低的氨基酸序列OMP81863.1作为预测模板,并使用Swiss-Model查询同源序列的蛋白质三级结构,从而预测得到LT_397蛋白三级结构。结果如图5所示:LT_397是单体,没有与其结合的配体;该蛋白质三级结构主要由2个螺旋结构并通过a螺旋和无规则的残基片段构成,并且通过β转角连接、折叠形成这种“背包形”的蛋白结构。
2.3 LT_397受逆境胁迫诱导转录分析
以LtActin基因作为内参基因,利用qRT-PCR分析不同侵染时问下可可毛色二孢对LT_397基因转录水平的影响。LT_397受到可可毛色二孢诱导上调表达,相对转录水平在24 h达到最高,是对照的2.85倍,之后转录水平逐渐下降(图6)。可可毛色二孢胁迫处理后效应子LT_397基因的相对转录水平均高于未胁迫处理样本(图7)。在营养吸收方面,氮元素的缺少诱导LT_397基因的相对转录水平上升明显,相对转录水平为18.86;氧化胁迫处理中,基因的相对转录水平随Hz 02浓度的增加呈上升趋势,10mmol/L Hz 02处理下基因的相对转录水平为15.20;热胁迫处理中,45℃高温处理的样本相对转录水平为3.98;在离子胁迫下,NaCl处理的样本中LT_397基因的相对转录水平是4.75,而LiCl处理下相对转录水平与对照水平相比,转录水平微上调,相对转录水平为2.30;酸碱胁迫处理发现:与适宜的pH值5~7相比,弱酸弱碱都会造成LT_397基因转录水平的上升,但碱性条件下基因的转录水平更高,相对转录水平为8.66;UV胁迫发现:紫外线照射时问越长,LT_397基因相对转录水平越高,UV照射10 min的LT_397基因相对转录水平为18.78。
2.4效应子LT_397可抑制颖壳伯克氏菌诱导的HR反应
效应子LT_397转入颖壳伯克氏菌感受态细胞后,在含有25mg/L庆大霉素的LB固体平板上挑取生长的单克隆转化子,进行菌液PCR验证。如图8所示:阳性转化子PCR扩增得到约1000bp大小片段,阴性对照pEDV空载体无假阳性的条带出现。将验证成功的转化子进行测序验证,也证明重组质粒pEDV-397构建成功。
植物病原真菌通常通过效应因子克服宿主免疫。颖壳伯克氏菌自身能够在本生烟上诱导很强的过敏性坏死反应。因此我们构建了pEDV-397载体,利用颖壳伯克氏菌三型分泌系统将效应子LT_397分泌到植物细胞内。结果如图9a所示,在本生烟上瞬时表达效应子LT_397,正常条件下生长3d后观察发现:瞬时表达效应子LT_397的叶片HR面积小于对照。选取10组处理样品测量HR反应直径,统计结果如图9b所示,在烟草叶片中,表达效应子LT_397的HR反应平均长度为0.55cm,而对照组平均长度为1.74cm,说明该效应子可抑制颖壳伯克氏菌诱导的HR反应。
3讨论
大量研究报道发现,植物病原真菌效应子通过多种途径抑制寄主植物的防卫反应,使病原真菌成功在寄主体内定殖、侵入和扩展。然而关于葡萄枝干病害的病原效应子功能的研究还未见报道。本试验克隆了可可毛色二孢特有的效应子LT_397基因,并利用生物信息学手段对该蛋白进行理化性质、二级结构和功能结构域预测分析,其编码的氨基酸N端具有一个典型的信号肽区段,含有11个Cys属于分泌蛋白。利用实时定量PCR技术分析LT_397基因在不同胁迫处理的转录模式发现:高温、H2Q2、UV处理均可诱导该基因转录水平的提高。
病原真菌效应子对植物免疫系统的调控作用,可通过烟草的细胞坏死反应进行快速检测。部分效应子在烟草的超表达可以引起烟草细胞坏死,另外的效应子可抑制不同激发子诱导的细胞坏死。本研究通过烟草毒力试验证实:效应子LT_397可抑制颖壳伯克氏菌诱导的HR反应,可能对可可毛色二孢的毒性有一定的促进作用。
可可毛色二孢是一种半活体营养型真菌,在侵染植物的早期需要从活体中吸取营养物质,而在侵染后期,则要杀死寄主植物,转换到死体营养,从死体植物组织中吸收营养物质。在可可毛色二孢中可能存在大量功能特异的效应子,它们可能在该病原侵染寄主过程中发挥重要作用。有关可可毛色二孢效应子的作用机制研究有待进一步通过基因编辑、蛋白互作等进行深入分析。通过探索LT_397等效应子的生物学功能以及作用机制研究,将有利于揭示可可毛色二孢的致病机制,进一步寻找和设计药物新靶点,为创建持久、高效的绿色防治技术提供理论基础。
