热河生物群窃蛋龙类的发现及研究进展*
2021-04-29裘锐汪筱林
裘锐,汪筱林
①中国科学院古脊椎动物与古人类研究所 脊椎动物演化与人类起源重点实验室,北京 100044;②中国科学院生物演化与环境卓越创新中心,北京 100044;③北京自然博物馆,北京 100050;④中国科学院大学 地球与行星科学学院,北京 100049
窃蛋龙类(Oviraptorosauria)是一类头部高度特化的手盗龙类,它们最显著的特征是除了原始类群之外头骨短而高,而且上下颌均没有牙齿[1]。第一块窃蛋龙类化石由美国自然历史博物馆中亚考察团于1923年发现于蒙古戈壁上白垩统地层,并在1924年由Osborn将其命名为嗜角窃蛋龙(Oviraptor philoceratops)[2]。窃蛋龙意为“偷蛋贼”,因为这具化石最初被发现时趴在一窝恐龙蛋附近,Osborn认为它可能是在偷吃其他恐龙的蛋。种本名“嗜角”意为“爱吃角龙类的”,但由于蛋中并无胚胎保存,Osborn自己也承认这样的推测不很可靠,他在论文中提到“这样的命名可能使我们完全混淆其本来的习性”[2]。近20年来,在蒙古和我国发现的多件窃蛋龙类及胚胎化石显示,窃蛋龙类并非偷蛋贼,而是具有趴伏在蛋窝上保护或孵化自己后代的习性[3-7]。除此之外,一些人将窃蛋龙类的吻部与现生食卵动物对比,发现现生食卵动物通常有较为宽大的牙齿,有时牙齿表面还比较粗糙,但是窃蛋龙的吻部是比较尖的,起到的作用更像切割而不是打破蛋壳[8]。
早期发现的窃蛋龙类恐龙并未被认为是单系群,这些恐龙被归入似鸟龙类(Ornithomimosauria)[2],或是原始鸟类[9-10]。直到1976年,蒙古古生物学家Barsbold发现这些恐龙都具有左右齿骨呈U形愈合、指爪近端关节面具有背唇突等共同特征,于是在虚骨龙类(Coelosauria)下建立窃蛋龙类[11]。1986年,美国古生物学家Gauthier根据窃蛋龙类具有近四边形的乌喙骨、后弓的尺骨、扩大的半月形腕骨、较细弱的第III指、坐骨一般不足耻骨长度的2/3、股骨第IV转子退化等,将其归入手盗龙类(Maniraptora),其系统发育位置与恐爪龙类(Deinonychosauria)和鸟类组成的单系群形成姐妹群[12]。但也有研究显示,镰刀龙类(Therizinosauroidea)和窃蛋龙类共同具有眶前孔变小、U形的齿骨联合、基蝶骨的基翼骨突短等特征,在一些系统发育结果中窃蛋龙类和镰刀龙类构成姐妹群,位于手盗龙类的基干位置[13-15]。还有些系统发育研究根据窃蛋龙类具有位置较高的鼻孔、缩短的鼻骨、扩大的顶骨、棒状的颧骨、外翼骨关节上颌骨而非颧骨、无齿的上下颌等特征,认为窃蛋龙类是与鸟类亲缘关系最近的恐龙,与鸟类形成姐妹群[16]。
早期研究对窃蛋龙类系统发育位置存在争议的主要原因是当时所有已知的窃蛋龙类都生存于晚白垩世,缺乏早期物种的化石记录。直到20世纪90年代之后,才在我国的辽宁西部热河生物群陆续发现早白垩世的窃蛋龙类[17-22]。尾羽龙科(Caudipteridae)[19]是热河生物群窃蛋龙类的代表之一,它们具有不同于晚白垩世窃蛋龙类的特征,有些标本还保存有非常精美的羽毛等软组织印痕[17-19],对研究窃蛋龙类早期演化和羽毛的起源与演化具有重要意义。
1 热河生物群尾羽龙科的发现历史及进展
尾羽龙科最早研究的物种是1998年命名的邹氏尾羽龙(Caudipteryx zoui,图1(a)),化石发现于辽宁北票四合屯下白垩统义县组。邹氏尾羽龙的模式标本前肢和尾部保存了精美的羽毛,特别是尾巴末端的羽毛呈扇形排列,与现代鸟类相似,故而得名[17]。在最早的研究中,邹氏尾羽龙被认为是手盗龙类中一个与鸟类亲缘关系很近的属种,而非窃蛋龙类[17]。有的研究甚至认为尾羽龙不是长羽毛的恐龙,而是次生失去飞行能力的鸟类,主要依据是牙齿基部收缩、肋骨上存在钩状突、缩短的尾部、髂骨上髋臼冠缺失等与原始鸟类共享,而这些是在当时发现的兽脚类恐龙中少见的骨骼特征[23]。
2000年,周忠和和汪筱林等研究了三块新的尾羽龙标本,并将其中一块发现于辽宁北票下白垩统义县组的标本命名为董氏尾羽龙(Caudipteryx dongi,图1(b))[18]。对新的尾羽龙标本研究显示:尾羽龙具有的一些骨骼特征,包括未愈合的胸骨、远端膨大的肩胛骨、肩胛骨和乌喙骨夹角大于90°、耻骨指向前腹部、坐骨闭突大、后肢各骨骼未愈合等,相比于原始鸟类,与其他兽脚类恐龙更为相似[18-19]。研究人员也发现,尾羽龙具有的一些原本认为与原始鸟类相似的特征后来也陆续发现于其他兽脚类恐龙[5,24]。周忠和和汪筱林据此认为尾羽龙属于窃蛋龙类,并建立尾羽龙科[19]。后续系统发育研究认为尾羽龙科位于窃蛋龙类的基干位置[14,16]。尾羽龙科的主要特征包括:前颌骨着生牙齿,而且牙齿弯曲;齿骨未愈合;尾椎数量仅25枚左右;第I掌骨长度短于第II掌骨的1/2;肠骨髋臼前突背腹高度较高;耻骨柄前后宽大于坐骨柄;第III跖骨近端呈窄跖型等。
2008年,另一块尾羽龙科标本发现于辽宁义县西二虎桥下白垩统九佛堂组,被研究命名为义县似尾羽龙(Similicaudipteryx yixianensis,图1(c))[21]。似尾羽龙与尾羽龙的主要区别在于最后几节尾椎愈合形成匕首状尾综骨、背椎具两个椎体下突、肩胛骨细长、髂骨较短等。尾羽龙最突出的特点是手部第III指极度退化,仅剩两根很短的指节,其指式为2-3-2,不同于其他大多数非鸟兽脚类恐龙的2-3-4。由于义县似尾羽龙没有保存手部骨骼,并不能确定该指式是否存在于似尾羽龙。

图1 尾羽龙科化石:(a)邹氏尾羽龙[17];(b)董氏尾羽龙 [54];(c)义县似尾羽龙[21]
包括尾羽龙科在内的一些原始窃蛋龙类的发现,对窃蛋龙类的原始特征进行了补充,对其系统演化位置的认识也逐渐清晰。现在系统发育结果普遍认为,窃蛋龙类是廓羽盗龙类(Pennaraptora)的基干类群,和副鸟类(Paraves)形成姐妹群[25-26](图2)。

图2 虚骨龙类(Coelosauria)的系统演化。可见窃蛋龙类(Oviraptorosauria)位于廓羽盗龙类(Pennaraptora)的基干位置,接近包括鸟翼类(Avialae)在内的副鸟类(Paraves)(修改自 [25])
2 尾羽龙科干戚刑天龙的发现及虚骨龙类的演化
2019年,《科学报告》(Scientific Reports)发表了我们对尾羽龙科新标本的研究,详细记述了在辽宁义县王家沟下白垩统义县组的尾羽龙科一新属种——干戚刑天龙(Xingtianosaurus ganqi,图3)[22]。干戚刑天龙虽然缺失头部和大部分的颈椎,但其较短的第I掌骨、缩短的尾部等特征显示该物种为尾羽龙科的一员。刑天龙与其他已知的尾羽龙科属种的主要区别包括背椎椎体侧凹接近椎体背缘、肩胛骨长度短于肱骨、尺骨长度与肱骨接近、桡腕骨角度较小、第I掌骨长度不足第II掌骨的40%等,系统发育结果显示干戚刑天龙为一原始的尾羽龙类。

图3 干戚刑天龙化石[22]
刑天龙的手部第III指虽然没有完整保存,但保存有第III指爪和部分指节,显示刑天龙的第III指并没有像尾羽龙的那样特化缩小。除此之外,刑天龙手指指节的韧带窝较小,而尾羽龙的手指指节的韧带窝极大。综合来看,刑天龙的手部特征更接近其他兽脚类恐龙而不同于尾羽龙。研究者以此为契机全面梳理了窃蛋龙类手部的演化,发现窃蛋龙类的手部演化非常复杂,各个支系都有各自不同的演化趋势(图4)。原始窃蛋龙类的指节I-1的近端关节面轻微凹陷,第II掌骨与第III掌骨长度接近,所有指节韧带窝较小,指节II-1与II-2长度之和大约是第II掌骨长度的1.1倍。尾羽龙科的进步属种,出现了第II指的指节伸长、韧带窝变大、第III指退化等特征变化。进步窃蛋龙类(新颌龙超科Caenagnathoidea),指节I-1的近端关节面明显凹陷。新颌龙科(Caenagnathidae)和尾羽龙出现趋同演化,指节II-1与II-2长度之和达到第II掌骨长度的1.5倍。除此之外,新颌龙科的第II掌骨长于第III掌骨。窃蛋龙科(Oviraptoridae)的河源龙亚科(Heyuanninae)第I指变得极其强壮,第II指的指节长度缩短,指节II-1与II-2长度之和短于第II掌骨。

图4 (a)虚骨龙类桡腕骨演化,可见越接近鸟类,桡腕骨角度越大;(b)窃蛋龙类手部的演化[22]
刑天龙的发现还为桡腕骨角度在虚骨龙类中的演化提供了新的信息。桡腕骨角度是桡腕骨与桡骨的关节面以及与半月形腕骨的关节面之间的夹角,该角度大小与手部和尺骨间活动角度相关。之前研究认为在兽脚类恐龙向鸟类演化过程中,桡腕骨角度大小总体呈上升趋势,从暴龙类(Tyrannosauroidae)的不足10°到原始鸟类的55°[27]。但也发现窃蛋龙类的桡腕骨角度异常增大,达到76°,不仅远大于其他兽脚类恐龙,甚至超过了原始鸟类。由于前人研究中窃蛋龙类的桡腕骨角度仅仅测量了尾羽龙,针对这一数据不足的问题,研究者测量了其他桡腕骨保存较好的窃蛋龙类,发现它们的桡腕骨角度都非常大,如哈格里芬龙(Hagryphus)为93°,可汗龙(Khaan)为62°,曲剑龙(Machairasaurus)为41°。但研究者注意到,这些测量数据要么来自于极度特化的类群(尾羽龙),要么来自晚白垩世进步的窃蛋龙类,不能很好地代表这一类群的早期演化趋势。刑天龙作为原始的、手部非特化的窃蛋龙类,其桡腕骨角度应该更接近这一类群演化初期的状态。干戚刑天龙的桡腕骨角度为39°,与虚骨龙类桡腕骨角度的整体演化趋势相吻合,证明了窃蛋龙类的桡腕骨角度初期其实比较小,在后来的演化中才逐渐增大,与鸟类趋同。
除了手部之外,刑天龙的胸骨也展现出和尾羽龙不一样的特征。尾羽龙的胸骨是两片椭圆形的骨头[18-19],而其他窃蛋龙类胸骨的前后边缘平,具有前侧突和侧突两个突起[28-30]。相比于尾羽龙,刑天龙的胸骨形态更接近于其他廓羽盗龙类恐龙。这一现象说明,尾羽龙独特的胸骨构造是独立演化而来的,并不代表窃蛋龙类胸骨的原始状态。
3 尾羽龙科的软组织保存及其研究进展
鸟类羽毛的演化一直是鸟类起源研究的重点方向之一。近20年来在我国辽宁西部、河北北部、内蒙古东南部发现的晚侏罗世燕辽生物群、早白垩世热河生物群发现的大量带羽毛的恐龙化石,其中保存了多种羽毛形态,为研究羽毛演化提供了非常重要的依据[17-18,24,31-33](图5(a))。目前认为鸟类羽毛最原始的状态是在美颌龙类(Compsognathidae)、暴龙类等身体表面覆盖的呈单根细丝的毛状结构[33-35]。羽毛演化的第二阶段是根部聚合且随后辐射发散的多分枝状,类似现代鸟类的绒毛,在部分镰刀龙类及驰龙类(Dromaeosauridae)的身上发现[36]。尾羽龙科的羽毛处于羽毛演化的第三阶段。从外形看,尾羽龙科的羽毛已经与现代鸟类非常相似,具有羽轴和羽片,但没有羽小枝和羽小钩存在的证据,而且羽轴左右两侧羽片是对称的(图5(b)),不像现代鸟类的具有空气动力学功能的羽毛是左右不对称的[17-18]。类似现代鸟类的、具有羽小枝和羽小钩的不对称飞羽则发现于与鸟类关系更近的驰龙类和伤齿龙类(Troodontidae)的一些属种[32,37]。

图5 (a)恐龙和鸟类羽毛的几种主要形态,从左至右分别为单根细丝状、多分支状、对称正羽状、不对称正羽状[36];(b)尾羽龙羽毛[17]
2010年,对似尾羽龙幼年个体的研究显示,似尾羽龙幼年个体身上附着的是绒毛,而另一块年龄大一岁的似尾羽龙标本则披覆正羽。这一研究显示尾羽龙科可能已经具有与现代鸟类相似的换毛行为,随个体成长发育,体表的羽毛会由绒毛变为正羽[38]。
羽毛起源的原因一直存在争议,鸟类羽毛最初出现的单根细丝状、多分枝状以及对称正羽都不可能用于飞行。角蛋白研究也显示恐龙羽毛的蛋白质以α-角蛋白为主,不像现代鸟类羽毛主要成分是能够提高柔韧性和强度的β-角蛋白,意味着恐龙羽毛尚缺乏足够强度以支持飞行[39],这说明羽毛最初起源肯定与飞行无关。近些年来,对恐龙羽毛颜色的研究取得较大进展[37,40-42]。研究显示,许多带羽毛恐龙的羽毛具有绚丽的彩虹光泽[37,42]。有些恐龙,比如近鸟龙,在头顶具有醒目的红色羽毛[41]。这些恐龙羽毛具有的色彩和光泽与今天鸟类求偶时炫耀的羽毛相似,推测羽毛的出现很可能是因性别炫耀的需求而产生的[43]。虽然在尾羽龙科标本尚未有颜色证据的发现,但也有一些暗示羽毛用于炫耀的证据存在。尾羽龙的羽毛虽然没有黑素体的研究,但其尾羽存在黑白相间的条纹[17]。对巴西早白垩世的羽毛化石研究显示,存在羽毛明暗色带交替的现象,其暗色区域具有黑素体,而浅色部分缺乏黑素体[44]。这一研究说明在尾羽龙活着的时候,其尾羽至少是明暗相间,具有白色条纹。这种具有花纹的羽毛很可能与现代鸟类的羽毛一样,起到性别展示的作用。
在尾羽龙的前肢肱骨近端与腕骨之间,保留有软组织痕迹,这一部分软组织与现代鸟类的翼膜所处位置非常相似[45]。翼膜对现代鸟类飞行时控制翅膀有很重要的意义,只有个别次生失去飞行能力的鸟类缺失翼膜。根据翼膜的存在,有一些研究推测,窃蛋龙类的原始物种是树栖动物,只是后来重新转移到了陆地生存[45-46]。但这一推测尚无化石证据支持。
4 尾羽龙科的胃石及食性研究
大多数兽脚类恐龙是食肉动物,具有锋利的牙齿和爪子,用来捕食恐龙或其他动物。但窃蛋龙类的牙齿完全退化,头骨变得短而高[11],对于其食性一直存在争议。尾羽龙科的发现为解决窃蛋龙类的食性问题提供了依据。现已发现多件尾羽龙科的标本的腹腔内保存有许多小石子(图6)[17-19],其形态和位置与现代鸟类和化石鸟类的胃石接近[47-49]。
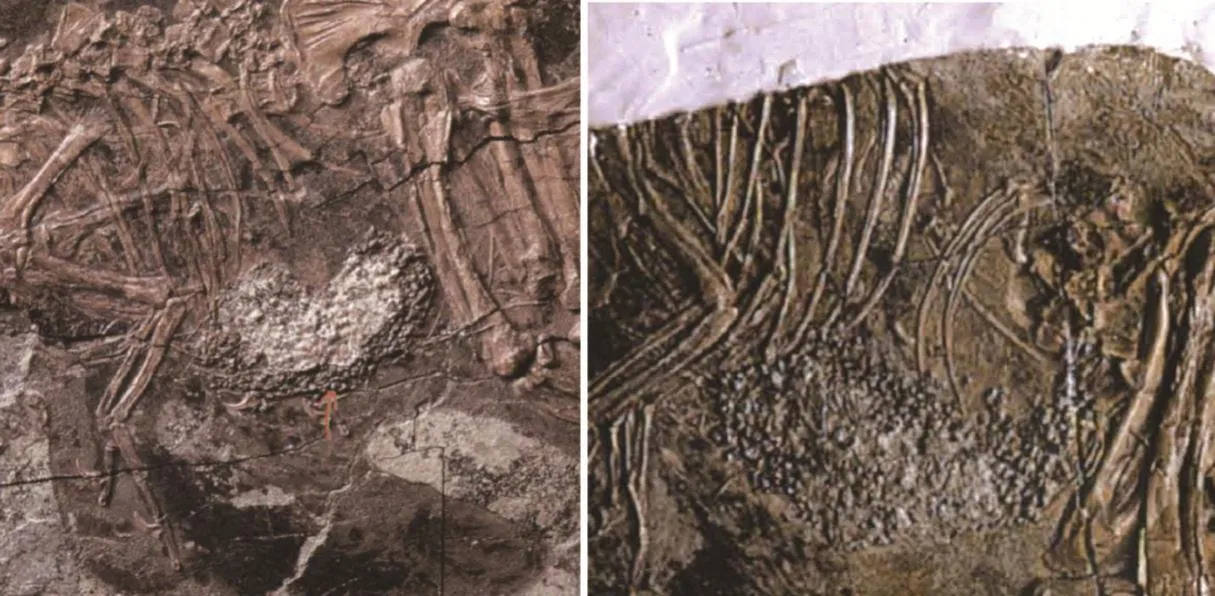
图6 尾羽龙标本腹腔内所含胃石[54]。左图:尾羽龙未定种;右图:董氏尾羽龙
胃石在现代植食性鸟类中尤其常见。现代鸟类由于牙齿退化,无法咀嚼坚韧的植物只能整个吞入,它们会主动吞食一些小石子并储藏在肌胃中,利用这些胃石来起到研磨食物的作用[50]。除此之外,胃石也有补充身体所需矿物质、清理胃壁等作用[48]。除植食性鸟类之外,现生和化石植食性爬行类也具有胃石[48,51]。肉食性鸟类或爬行动物偶尔也有胃石发现,它们的胃石一般数量少,大小不一,而且位置接近肠道而非胃部,一般认为是吃食物时偶然吞入所致[52-53]。尾羽龙科标本保存的胃石数量非常多,排列紧密,保存位置紧贴胸骨之后,直径较小,一般不到4.5 mm,与植食性鸟类的胃石接近。这一发现显示,尾羽龙科很可能是一类植食性的恐龙。进一步的研究发现,包括尾羽龙科在内的多种原始窃蛋龙类的牙齿退化,牙齿仅局限于上下颌的最前部,或仅存于前颌骨[17,19-20],而且牙齿形态特化,不同于其他兽脚类恐龙尖锐、后弯的匕首形牙齿,原始窃蛋龙类的最前部的牙齿较钝并增大、扁平,形态接近哺乳动物的门齿[17,19-20],而且这种牙齿适合切断植物。结合胃石和牙齿形态研究,包括尾羽龙科在内的原始窃蛋龙类很可能是植食性恐龙,而窃蛋龙类可能是一类由肉食性转变为植食性的兽脚类恐龙。
综上所述,近些年发现的包括尾羽龙科在内的窃蛋龙类,丰富了我们对窃蛋龙类早期演化、窃蛋龙类的系统发育位置、鸟类羽毛的早期演化及生活习性等方面的认识。但对窃蛋龙类是否存在树栖原始物种、早期窃蛋龙类骨骼特征演化的功能意义等方面尚存疑问。还有必要针对现有化石进行更细致的观察比较,并与胚胎发育学等学科交叉,结合野外工作可能发现的新化石,对这些问题进行研究。
