叶片扁平化的形成机制
2021-04-29汪颖焦雨铃
汪颖,焦雨铃
①中国科学院大学 生命科学学院,北京 100049;②中国科学院遗传与发育生物学研究所 植物基因组学国家重点实验室/国家植物基因研究中心(北京),中国科学院种子创新研究院,北京 100101;③中国科学院大学 现代农业科学学院,北京 100049
自然界中叶片的形态千变万化,有单叶和复叶之分:单叶在一个叶柄上只生有一个叶片;复叶在一个叶柄上生有两个或更多叶片。相对简单的单叶的外形还有很多类型。一位植物学家可以列举出圆卵形、披针形等十余种单叶叶型。单叶的叶缘可以具有或深或浅的缺刻,例如枫叶。深裂单叶与复叶有时已经难以区分。复叶的形态更为多变。尽管形态如此多变,它们大多具有扁平化的特征,因而被归为叶片[1]。
扁平化叶片的出现是进化上的一个重要事件[2-3]。对化石研究的结果表明,植物登陆于4.2亿年前的志留纪晚期。最早的陆生植物只有分枝,没有叶片,光合作用是由分枝来完成的。在植物登陆后最初的约4 000万年间,陆地上出现了几米高的大树,但是直到距今3.6~4.0亿年前的泥盆纪晚期叶片才出现。叶片的出现伴随着大气中90%二氧化碳的固定,极大地改变了地球生态系统,为其他物种的出现铺平了道路。
随着叶片扁平化的形成,叶片两面产生并呈现差异。如果细心观察过植物的叶片,你应该可以轻易分辨出叶片的正反两面:叶片的向阳面(也叫近轴面,即朝向茎的那一面)比较光滑,有比较厚的蜡质层,并且颜色更翠绿一些;而相对地,向地面(远轴面)就要粗糙一些,角质层也薄一些(图1)。

图1 叶片的正反两面往往具有很大的差别
这其实是植物针对自己的生存和生理功能演化出的特征,对于植物的光合作用及蒸腾作用很有好处。叶片近轴面主要的功能是吸收阳光中的能量,进行光反应,因此叶肉细胞拥有比较多的叶绿素;相对来说,叶片的远轴面就没有那么多的叶绿素了,而是主要进行光合所需的气体交换,因此叶片背面分布的防止水分散失的角质层自然也薄一些,颜色也要淡一些。事实上,光照本身能促进叶绿素的合成,在光照不足的地方生长的植物就会因为缺乏叶绿素而变得黄黄的。
1 叶片发育过程中扁平化的逐步建立
叶片是由茎尖分生组织产生的。茎尖分生组织的中心维持着一团干细胞,在周边产生器官。在种子植物开花前,茎尖分生组织周边不断产生叶片原基(图2)。叶片原基最初大致为半圆形,但各个方向的生长速率显著不同。最终形成的叶片具有基-顶轴、中-边轴和近-远轴三个轴向。其中基-顶轴由叶片基部指向叶尖;中-边轴从叶片中线指向叶缘;近-远轴即前面提及的“正反面”,靠近茎尖分生组织一侧为近轴面。
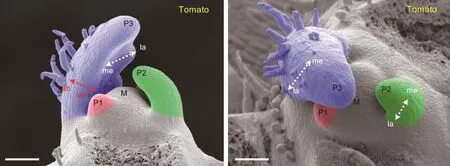
图2 番茄茎尖分生组织周围产生叶片原基(修改自[4])。最小的原基称为 P1,之前发生的为 P2、P3……。M标示茎尖分生组织中心。ad-ab标示近-远轴,me-la 标示中-边轴。标尺为 50 μm
叶原基形成后,三个轴向的生长速率有显著不同。叶片在近-远轴方向的生长速率远小于中-边轴和基-顶轴。以模式植物拟南芥(Arabidopsis thaliana)为例(图3),近-远轴向的细胞层数从很早开始就维持在6~7层,直至叶片成熟。中-边轴和基-顶轴则快速延长,形成叶片。由此可见,叶原基定向生长模式的建立是扁平化叶片形成的关键。

图3 拟南芥初生叶片原基的横切面比较。叶片标记如图2。注意叶片近-远轴生长有限,远小于中-边轴生长速率。标尺为 50 μm
2 叶片扁平化依赖近-远轴极性
叶片生长的三个轴向几乎同时建立,那么它们之间是否有依赖关系呢?一种检测方法是打破一个轴向的建立,看看其他轴向是否受影响。
分子遗传学技术是研究生命过程的有力手段。基因可能自发突变,而人为处理还可以加速突变。基因突变后如果一个发育过程出现异常,那么我们就可推断突变的基因调控这个发育过程。通过分子遗传学,我们再找出突变的基因,这样就能把基因和发育过程对应上。这就像拆汽车:当汽车的某个零件被卸掉,车灯就不亮了,那么这个零件就是负责车灯正常工作的。
利用分子遗传学技术,科学家们发现了很多叶片不能正常扁平化生长的突变体,其中最早研究的是金鱼草(Antirrhinum majus L.)中的一个自发突变体 phantastica (phan)[5]。phan突变体的叶片有一部分异常,呈现变窄、变为喇叭状或棒状表型。对这些异常叶片的细胞形态和解剖结构进一步检查,研究者发现叶片趋于远轴面化,即叶片的表皮、叶肉、维管束中近轴面区域细胞部分或全部显现出远轴面细胞的特征。与此同时,叶片不能扁平生长,即中-边轴向发育被抑制。基于此,科学家提出中-边轴向的建立依赖于近-远轴向的正确建立。
更多的突变体研究也支持这个观点。拟南芥中一系列影响近轴面或远轴面发育的突变体都趋于叶片变窄及形成喇叭叶或棒状叶,说明叶片扁平生长受到影响[1]。然而,烟草(Nicotiana sylvestris)仅形成中脉区域,无法产生宽大叶片的lamina1(lam1)突变体仍能维持基本的近-远轴向,说明叶片近-远轴的建立并不严格依赖中-边轴向发育[6]。
3 基因表达界定叶片各区域
如前所述,叶片的不同区域具有不同的形态特征和生理、生化功能。区域特异的基因导致了各区域的差异,其中最为重要的有近轴面表达的HD-ZIP III类转录因子、远轴面表达的KANADI(KAN)转录因子以及中间区表达的WOX类转录因子。在拟南芥中,HD-ZIP III类转录因子表达于分生组织区,并在叶原基的近轴面有更强的表达[1]。编码HD-ZIPIII类转录因子的mRNA均被小RNA miR165/166降解[7]。这些小RNA表达在远轴面的表皮,并扩散至整个远轴面区域[8-9]。拟南芥中冗余的KAN 转录因子环绕茎尖分生组织表达,并在新形成的叶原基中处于远轴面区域。活体成像表明,这两类基因的表达在叶原基形成前就已经出现,形成了“预置模式”,叶原基跨两个区域形成[10-11]。伴随叶原基的形成,编码WOX转录因子的PRESSED FLOWER(PRS)在近、远轴区域之间开始表达,界定了中间区。
在叶片发育早期,近轴面、中间区和远轴面三个区域一直维持。近轴面和中间区有部分重叠,但二者和远轴面基本无重叠。中间区是叶片展开的关键区域。烟草的lam1突变体正是中间区表达的WOX基因发生了突变[12]。单、双子叶植物中中间区表达的WOX基因突变均会影响叶片的形成,说明该机制高度保守。因物种而异,中间区表达的WOX基因或集中在叶片外缘,或遍布整个叶片的中间区域。WOX基因促进细胞分裂,为叶片生长提供新的细胞。在叶片成熟后,WOX基因表达终止,叶片不再长大[13]。
在近轴面、远轴面基因缺失或异位激活的突变体中,WOX的表达区域消失或不再出现于叶缘。这从分子水平描述了叶片的中-边轴发育依赖于近-远轴的建立,同时也提出了问题:WOX的表达如何被近、远轴面的基因调控?
4 经典的植物“外科”实验
在分子遗传学技术出现之前,发育生物学家已经发现了一些调控叶片展开和近-远轴极性建立的线索。我们先从一个经典实验开始。
20世纪四五十年代,曼彻斯特大学研究生Ian Sussex以土豆为实验材料,研究植物茎尖与叶片的关系。之所以选择土豆,是因为它不仅容易买到,而且能不断生芽,人们可以方便地削下带着芽的土豆块,修成合适的形状放在显微镜下观察。
Sussex用手术刀把即将发生的叶片原基和茎尖轻轻划开一个切口,但是让叶片的其他部位与整体相连(图4)。几天后,他发现那些在近轴面与茎尖分开的叶片无法侧向展开,形成棒状叶片。通过对细胞形态进行细致观察,他发现这些棒状叶片的近-远轴极性发育不正常,叶片各个方向都和远轴面相似。此外也有些叶片成为“喇叭叶”,其顶端像喇叭一样是圆锥形的,喇叭口以下与棒状叶类似,没有极性[14-15]。
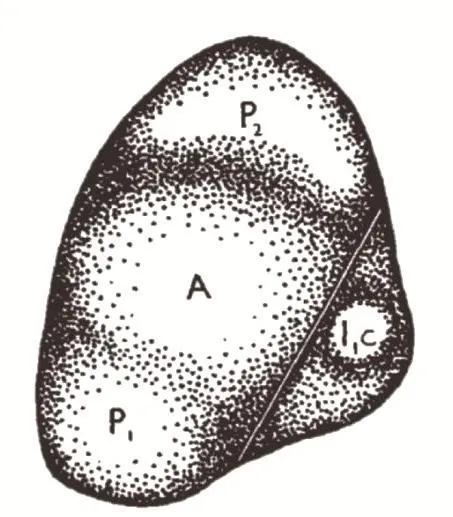
图4 Sussex在即将发生的叶片原基(I1c)和茎尖(A)之间切开一个小口,但是底端仍与整体相连[15]
Sussex由此推测:茎尖的干细胞产生一个信号,指导叶片近轴面形成;当“手术”切割阻断这个信号时,默认的远轴面发育程序启动,导致叶片只有远轴面发育。该实验最后被写入了发育生物学的教科书,而这个信号也因此被称为“Sussex信号”。
5 一切都指向生长素
Sussex实验似乎很好地解释了极性的信号来源,可是对于它到底是由什么物质介导,甚至它是否真的存在,一直众说纷纭。近半个世纪后,我国许智宏先生课题组的一项研究打破了这一领域的沉寂。该团队在组织培养的白菜、烟草等多种植物中都发现,加入了生长素的运输抑制剂能够诱发喇叭叶和棒状叶产生。这说明生长素和它的运输与叶片极性发育有关[16]。
生长素是最早被发现的一类植物激素,其中最重要的化学物质为3-吲哚乙酸(indole-3-acetic acid,IAA)。这种小分子可以由色氨酸合成得到,虽然对人和其他动物没有什么作用,却对各种植物的发育至关重要,根生长、维管束分化、顶端优势、单性花形成、向光性和向地性等都与它相关。生长素在植物体内的分布很大程度上取决于定向运输,而定向运输则依赖于一类称为PIN的蛋白(图5)。正是PIN蛋白介导的生长素运输产生了叶片极性发育信号。

图5 在植物体内,生长素通过PIN蛋白(绿色)从组织的一端传递向另一端(图片来源:The Plant Cell / Teaching Tools in Plant Biology)
信号分子生长素通过主动运输在茎尖表皮动态分布(图6(a))。叶原基形成的位置就是生长素汇聚的高点。在原基隆起后,原基的近轴面基部与茎尖分生组织间形成了边界区,生长素从边界区向两侧分流。我们发现,拟南芥中叶原基近轴面区短暂维持低生长素状态,而中间区生长素水平很高。这个生长素的差异分布依赖于PIN1介导的主动运输。在番茄中局部阻断从边界区运向茎尖分生组织的生长素流能够模拟手术切割,获得不能扁平化生长的棒状叶、喇叭叶[17]。在近轴面局部施加生长素也会得到和Sussex实验类似的结果。由此看来,生长素从叶原基向茎尖分生组织的运输介导了这个信号,只是运输方向和Sussex的推测正好相反。
在叶原基两侧的叶缘处,周边的PIN1均指向叶原基。黄海老师课题组发现,在叶缘两侧同时切口也会导致和Sussex实验类似的结果[18]。叶缘正是生长素最为富集的地方。叶原基两侧的切口很可能也是由于阻断了生长素的定向运输,从而影响叶原基内部的生长素分布。
前面提到,中-边轴生长依赖于近-远轴的建立,而正是生长素分布和信号介导了这个调控过程。生长素激活受体,释放下游转录因子MONOPTEROS(MP)激活转录。我们发现,MP在叶片的近轴面和中间区均有分布,但在远轴面无表达[19]。PIN1介导的生长素运输导致生长素在中间区富集,而近轴面短暂维持了低生长素环境。在生长素存在的情况下,MP可以直接激活界定中间区的 WOX 基因表达。由于生长素和MP分布的差异,WOX只在二者的交集也就是中间区表达,促进叶缘生长(图6(b))。

图6 茎尖生长素运输方向和生长素介导近-远轴界定中间区。(a)茎尖生长素的运输方向和分布。P1标示第一个叶原基,I1标示下一个未形成的叶原基。P1、I1周边的虚线分别标示 Sussex式手术切割和两侧手术切割位置。(b)叶原基纵切显示三个主要区域及生长素分布和信号分子分布如何决定中间区WOX基因表达
6 应力反馈维持扁平化生长
基因的表达如何调控三维形态的变化呢?植物细胞的生长是细胞内的膨压与细胞壁约束相互拮抗的结果。当膨压一定时,细胞壁约束弱则细胞生长快。我们发现早期叶原基的细胞壁硬度在近轴面、中间区和远轴面存在差异且随着发育快速变化[20]。同时,细胞壁组分果胶的甲酰酯化程度也一并变化,可能是影响细胞壁硬度的原因。这些力学属性的差异能够解释早期叶原基由接近辐射对称向两侧对称的转变。
此后,叶片维持高度定向的生长,在近-远轴向几乎没有加厚,是扁平叶片形成的关键(图3)。这种高度定向的生长和细胞分裂是如何实现的呢?中法合作的研究发现,叶原基发育早期的细胞生长也是高度定向的,以中-边轴向为主[4]。细胞的膨压没有方向性,因此决定生长的是细胞壁约束的各向异性。纤维素丝在叶原基细胞中高度定向排列,与生长方向垂直。纤维素的合成是由沿着微管运动的纤维素酶复合体完成的[21]。细胞中微管的排布与纤维素一致,以近-远轴向为主。力学计算表明,两侧对称的叶片原基中近-远轴向应力高于其他方向。微管的排布往往被应力所决定,沿主应力方向排布。这一系列分子水平的作用链组成了一个正反馈环:初步出现扁平化的叶原基中主应力沿近-远轴方向,导致微管沿此方向排布;微管指导纤维素丝的合成,使纤维素丝同向排布;纤维素丝的定向排布使细胞无法在近-远轴向进一步生长,而是沿着与之垂直的平面生长,使原基更为扁平化;扁平化的原基中主应力依旧维持近-远轴方向。以上反馈环的不断重复加剧了叶片沿中-边轴和基-顶轴的扁平化生长(图7)。

图7 应力反馈使叶片维持扁平化生长[4]
综上所述,中间区WOX基因和近、远轴面基因的表达使叶片原基从最初接近辐射对称的隆起变形为两侧对称,而应力反馈进一步维持高度方向性的生长使叶片展开。
7 叶片起源的应力反馈学说
叶片在进化过程中如何形成,一直是科学家们所热衷的话题。最初的植物只有不断分枝的枝条,由嫩枝进行光合作用。根据挖掘出的植物化石,考古学家 Walter Zimmermann 提出了顶枝学说(telome theory)[22]。顶枝指二叉分枝轴的顶端部分,可以分为不育顶枝(叶状枝)和能育顶枝(孢子囊)。顶枝学说假设了三步形态特化,认为顶枝通过这些特化过程进化为叶片(图8)。这包括邻近顶枝变为不等长度的“越顶”、邻近顶枝集中到同一平面的“扁化”,以及邻近顶枝间形成薄壁的“蹼化”组织。然而这些假设的形态特化很难和现存的发育过程对应上,更缺少分子证据的支持。

图8 顶枝学说假设的主要形态特化步骤:越顶、扁化和蹼化(修改自[1])
在应力反馈调控叶片扁平化研究的基础上,我们提出了解释叶片进化的“应力反馈学说”[23]。三维计算模拟表明,应力反馈不仅能使扁平化器官维持扁平化生长,还能使枝、根等辐射对称器官的生长以长轴方向为主[4]。应力反馈机制很可能是保守的机制,作用于各种植物器官类型。早期登陆的植物产生辐射对称原基,其在应力反馈作用下沿着长轴生长,逐步成为枝条。在进化过程中,原基中基因表达的差异性产生,导致原本辐射对称的原基呈现初步的扁平化,即两侧对称。两侧对称一旦产生,就被应力反馈放大,通过持续的扁平化生长形成宽大的叶片(图9)。随着地球环境的变迁和气孔等其他自身结构的出现,当扁平叶片有更好的适应性时,就被选择并保留,替代了原初仅有枝条的早期陆生植物。
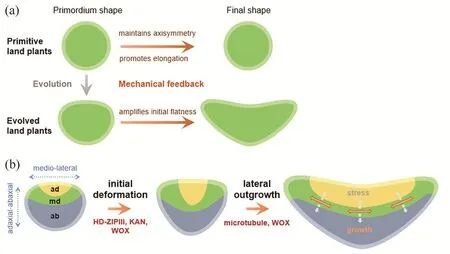
图9 叶片起源的应力反馈学说。(a)扁平叶片进化的主要步骤;(b)叶原基由初始形变产生初步扁平化形态,再由应力反馈维持扁平化生长形成叶片(修改自[23])
叶片的扁平化是一个基本的自然现象,在一代又一代植物发育生物学家的努力下,它的形成机制逐步被揭示。生物力学、数学模拟等新的研究方向极大拓展了我们的思路,带来了新的学说。随着生命科学和其他学科的突破,我们相信在不远的将来,对叶片扁平化的形成机理的理解会有进一步的加深和完善。
