MicroRNA-181d-5p通过PTEN参与睾丸支持细胞间紧密连接损伤*
2021-04-28张丽虹赵小贞
张丽虹 王 锋 赵小贞 王 玮
(福建医科大学人体解剖学系,临床应用解剖学研究室,脑老化与神经变性疾病福建省高校重点实验室,福州 350122)
MicroRNA 是真核生物体内一种内源性小分子单链RNA,通过与目标mRNA的3'UTR区不完全互补结合形成沉默复合物,抑制靶基因表达。睾丸内紧密连接位于生精小管基底部的支持细胞之间,是血-睾屏障(blood-testis barrier,BTB)最主要的组成部分,主要由3种跨膜蛋白(occludin、claudin、JAM)和外周胞质蛋白(zonula occluden,ZO)构成,最终锚定于细胞内肌动蛋白(actin)骨架系统[1]。研究显示,能够引起肌动蛋白骨架系统紊乱的因素同样会造成紧密连接损伤[2-4],例如PI3K/Akt信号通路的激活[5-13]及肌动蛋白相关蛋白2/3(actin-related-proteins 2/3,Arp2/3)非正常地活化及转移等[14]。黏着斑激酶(focal adhesion kinase,FAK)是一种非受体型酪氨酸激酶,可与紧密连接处的occludin 或ZO-1 等蛋白复合物结合[6,15],并通过转变其不同位点的磷酸化形式参与紧密连接调控[16-19],影响分枝状肌动蛋白骨架系统的成核及聚合过程[20-22]。人第10号染色体缺失的磷酸酶和张力蛋白同源的基因(phosphatase and tensin homology deleted from chromosometen,PTEN)是一种具有双重特异磷酸酶活性的抑癌基因,其编码的蛋白既可以作为脂质磷酸酶抑制PI3K/Akt信号通路激活,又可以作为蛋白磷酸酶使FAK的第397位酪氨酸发生去磷酸化,降低p-FAK-Tyr397水平[23]。本研究主要探讨miR-181d-5p 是否可以与PTEN mRNA的3'UTR区相互作用,并降低其表达,通过升高p-FAK-Tyr397水平以及激活PI3K/Akt信号通路影响肌动蛋白骨架系统稳态,造成紧密连接损伤。
1 材料和方法
1.1 主要试剂
DMEN/F-12、PBS(HyClone公司);胎牛血清(BI公司);PTEN、FAK、p-FAK-Tyr397、Akt、p-Akt-Thr308、β-actin兔抗鼠多克隆抗体(Cell Signaling公司);Lipofectamine 2000试 剂(Invitrogen公司);Opti-MEMⅠ、0.25%Trypsin-EDTA、DMSO、Penicillin-Streptomycin(Gibco公司);Dual-Luciferase Report Assay试剂盒(Promega公司)。
1.2 双荧光素酶报告基因分析实验
1.2.1 质粒构建 本实验所用质粒购买于吉凯基因公司,分别为miR-181d-5p NC空白对照质粒、miR-181d-5p过表达质粒、重组PTEN mRNA 3'UTR野生型荧光素酶报告质粒(PTEN-3'UTR-wt)、重组PTEN mRNA 3'UTR 突变型荧光素酶报告质粒(PTEN-3'UTR-mut)、内参海肾荧光素酶报告质粒。
1.2.2 细胞培养及分组 293T细胞采用常规细胞完全培养基培养,条件为37℃、5%CO2及饱和湿度。选对数生长期细胞进行实验。设置4个实验组,分别为PTEN-3'UTR-wt+miR-181d-5p NC组、PTEN-3'UTR-wt+miR-181d-5p组、PTEN-3'UTRmut+miR-181d-5p NC组、PTEN-3'UTR-mut+ miR-181d-5p组。每组设6个复孔,各组质粒转染48 h后,进行荧光检测,具体操作过程按照双荧光素酶报告基因分析系统发光试剂盒说明书进行。
同时在293T细胞上采用质粒单纯过表达miR-181d-5p,设置2个实验组,分别为293T+miR-181d-5p NC空白对照组(293T+miR NC组)和293T+miR-181d-5p过表达实验组(293T+miR组),每组设6个复孔。
1.3 TM4细胞过表达miR-181d-5p及细胞间跨膜电阻(transepithelial electrical resistance,TER)值测定
1.3.1 MicroRNA mimic构建由于TM4细胞对质粒的转染效率不高,因此miR-181d-5p过表达采用microRNA mimic(吉玛基因生物公司)进行,分别设为miR-181d-5p NC空白对照mimic 和miR-181d-5p过表达mimic。
1.3.2 细胞培养及分组 小鼠睾丸支持细胞TM4细胞购买于ATCC公司,采用常规细胞完全培养基培养,条件为37℃、5%CO2及饱和湿度,选择对数生长期细胞进行实验。设置2个实验组,分别为TM4+miR-181d-5p NC空白对照组(TM4+miR NC组)和TM4+miR-181d-5p过表达实验组(TM4+miR组),每组设6个复孔。各组mimic转染48 h后,进行相关信号分子和TER值测定。
1.3.3 TER值测定 使用Millicell ERS 电阻仪测定TM4细胞间TER值,在24孔板中放入插入小室测定,每组设6个复孔,具体操作过程参照仪器使用说明书,测定的单位面积电阻值的计算方法为:单位面积电阻值(Ωcm2)=电阻值(Ω)×有效跨膜面积(cm2)(24孔板的有效跨膜面积值为0.6 cm2)。
1.4 免疫印迹检测
24孔板转染后48 h,采用细胞裂解液提取总蛋白,BCA法测定总蛋白浓度。制备SDS 聚丙烯酰胺凝胶,并进行电泳分离,每孔上样量为20 μL,总蛋白上样量约为40 µg。转膜及脱脂奶粉封闭1 h后,采用一抗4℃孵育过夜(PTEN、FAK、p-FAK-Tyr397、β-actin稀释倍数为1:500,Akt和p-Akt-Thr308为1∶200),HRP标记的二抗(1∶2 000)室温孵育1 h,洗膜,ECL发光,分子生物学发光仪拍摄,Image J 软件计算条带灰度值,并计算各目的蛋白的相对表达量。
1.5 统计学处理
采用SPSS 21.0 软件进行统计学分析,符合正态分布的计量资料采用±s表示,采用单因素ANOVA 分析及Tukey 法进行多重比较,P<0.05为差异具有统计学意义。
2 结果
2.1 MiR-181d-5p可以与PTEN mRNA 3'UTR区结合,并降低其表达
双荧光素酶报告基因分析显示,质粒转染293T细胞的效率为60%~70%(图1A~D)。PTEN-3'UTR-wt+miR-181d-5p组的相对荧光值比率低于其他组(P<0.05)(图1E、F),证明miR-181d-5p可以与PTEN mRNA 3'UTR区结合。免疫印迹检测结果显示,293T细胞过表达miR-181d-5p后(图2A、B),PTEN蛋白表达降低(P<0.05)(图2C)。

图1 双荧光素酶报告基因分析各组质粒转染效率,标尺=100 μm
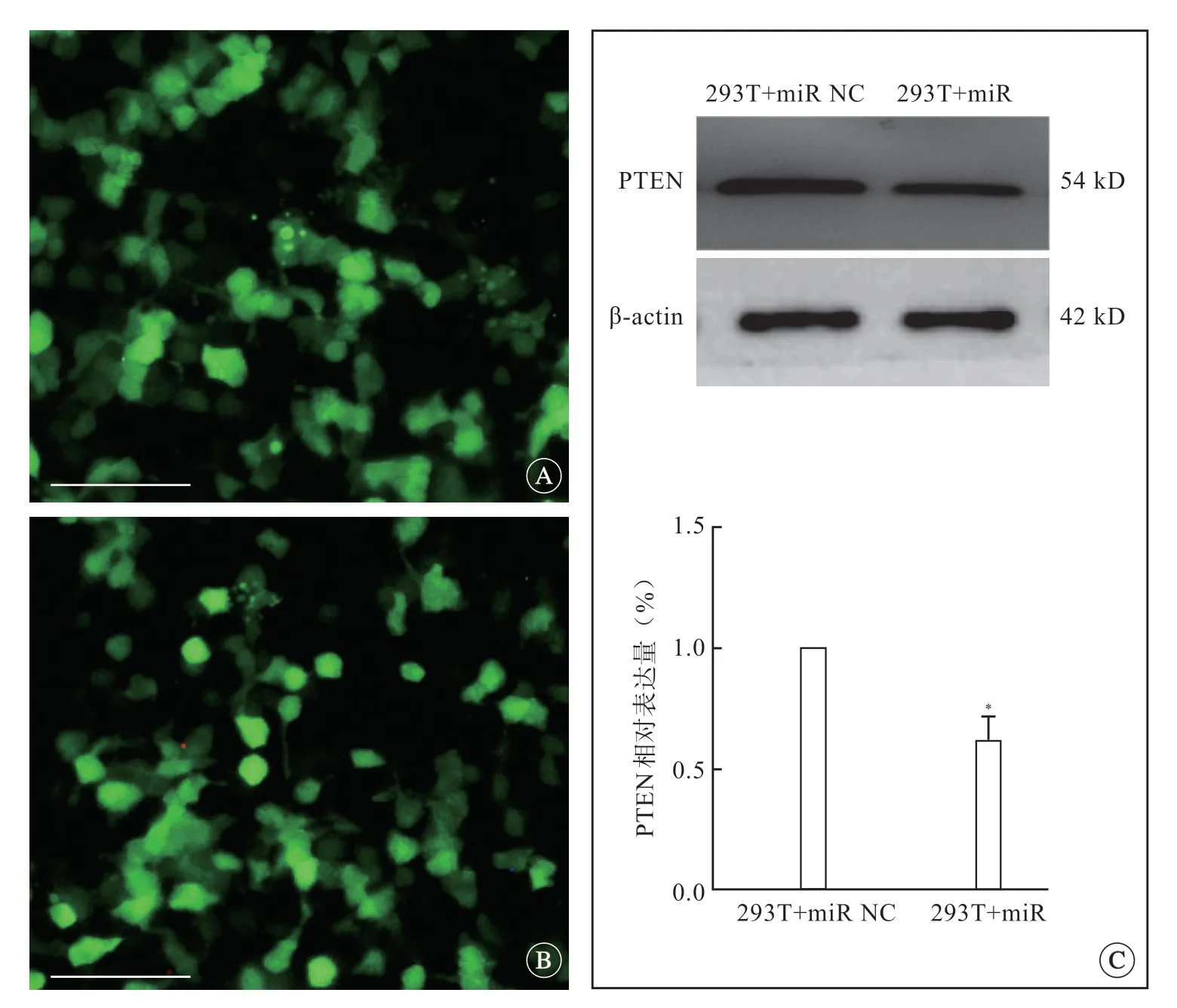
图 2 各组在293T细胞过表达miR-181d-5p 效率及PTEN蛋白表达,标尺=100 μm
2.2 TM4细胞过表达miR-181d-5p 引起TER值降低
结果显示,microRNA mimic 转染TM4细胞的效率为30%~35%(图3A、B)。TM4细胞过表达miR-181d-5p后PTEN蛋白水平降低,p-Akt-Thr308和p-FAK-Tyr397水平升高(P<0.05)(图3C、D),但Akt 及FAK表达水平没有变化。TM4细胞间相对TER值(0.70±0.15)降低。
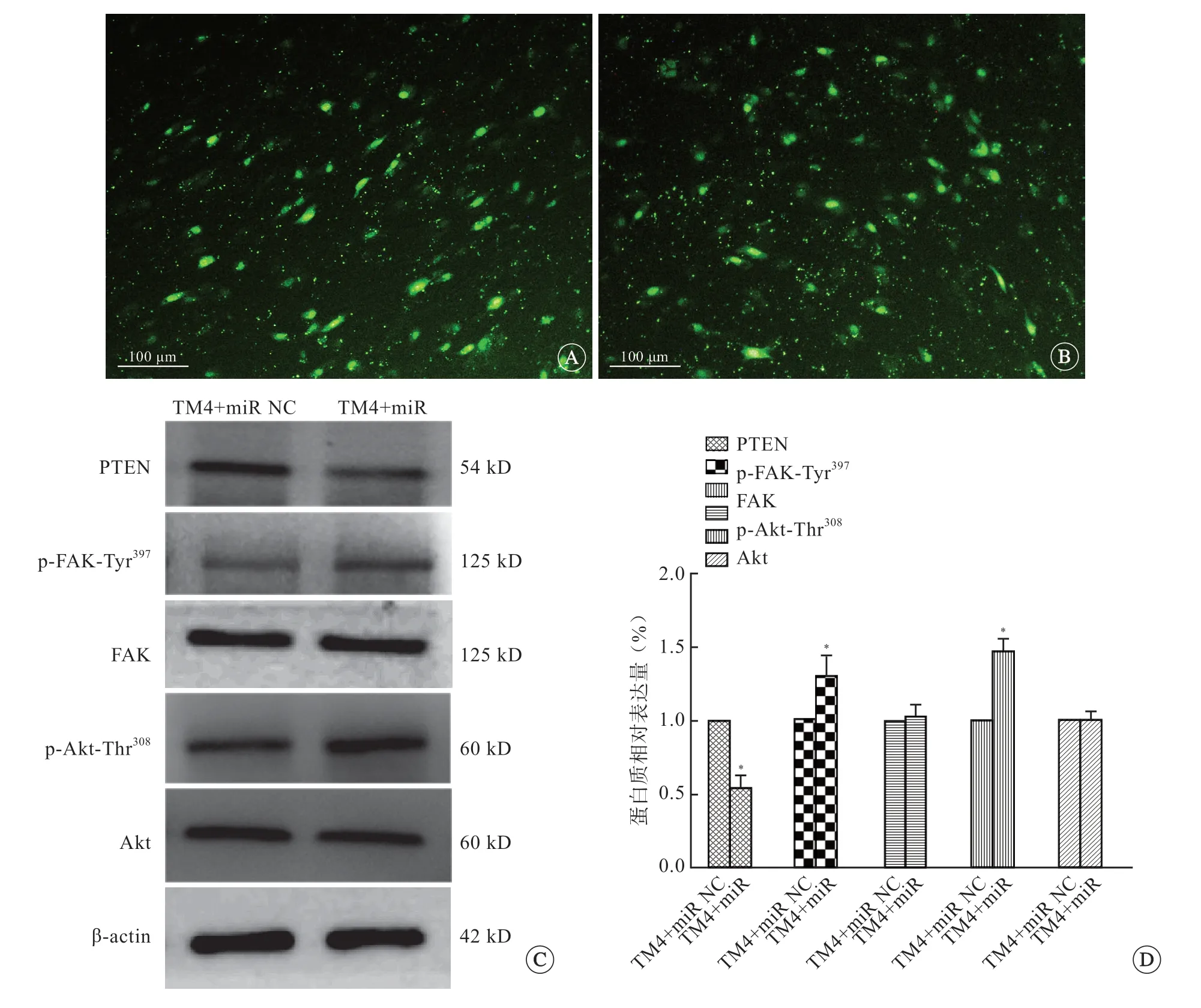
图3 各组小鼠睾丸支持细胞TM4细胞过表达miR-181d-5p 效率及各蛋白表达,标尺=100 μm
3 讨论
3.1 MiR181d-5p通过与PTEN mRNA 3'UTR结合降低PTEN蛋白表达
近年来研究显示,microRNA 无论是在生理还是病理条件下均发挥重要作用,不仅参与众多生物学过程,同时与疾病的发生和发展密切相关[24-25]。MicroRNA可通过与mRNA的3'UTR区不完全互补结合,进一步诱导mRNA的降解、切割及翻译抑制等过程,抑制靶基因的表达。本研究主要基于前期精索静脉曲张动物模型的研究结果开展,通过对睾丸组织进行microRNA测序、筛选及验证实验,显示精索静脉曲张会引起左侧睾丸组织内miR-181d-5p表达升高以及紧密连接结构和功能的的损伤[26]。通过查阅相关网站(http://www.targetscan.org/vert_71/)发现,PTEN mRNA的3'UTR区存在3个潜在可以与miR-181d-5p结合的位点(核苷酸序列为TGAATGT,分别为2261~2267位、2468~2474位和2675~2681位)。通过生物信息学分析,推测精索静脉曲张引起的miR-181d-5p表达水平升高,可能参与了睾丸内紧密连接损伤过程,并可能是通过降低PTEN的表达实现,但具体机制尚未明确。因此,设计本实验的目的是验证miR-181d-5p 是否可以与PTEN mRNA的3'UTR区结合,并降低其表达,通过PTEN 参与睾丸支持细胞间紧密连接的损伤过程,试图从microRNA层面阐述紧密连接动力学调控及损伤机制。本研究结果证实miR-181d-5p可以与PTEN mRNA的3'UTR区结合,且293T细胞过表达miR-181d-5p后可引起PTEN蛋白表达降低。
3.2 PTEN 水平降低造成睾丸支持细胞间紧密连接损伤的机制
PTEN 是一种具有双重特异磷酸酶活性的抑癌基因,与肿瘤的发生和发展密切相关。在本实验中,miR-181d-5p通过降低PTEN表达参与紧密连接损伤的机制可能主要涉及以下两个方面。
首先,PTEN作为脂质磷酸酶可以抑制PI3K/Akt信号通路的激活,它可使该信号通路的第二信使phosphatidylinositol(3,4,5)-trisphosphate(PIP3)发生去磷酸化转变为phosphatidylinositol(4,5)-bisphosphate(PIP2),从而抑制下游信号分子激活。PI3K/Akt信号通路的激活可通过扰乱肌动蛋白骨架系统稳态造成紧密连接损伤,而给予PI3K/Akt信号通路阻断剂,能够减轻由生殖毒性物质或其他理化因素造成的紧密连接损伤[5-13]。基于本实验结果,推测由miR-181d-5p过表达引起的PTEN表达水平降低,进一步激活PI3K/Akt信号通路,参与紧密连接损伤过程。此外,研究显示PIP2在细胞膜上的含量需>5%,才可启动正常地由WASP(Wiskott-Aldrich syndrome protein)家族蛋白和Arp2/3 介导的肌动蛋白成核和聚合过程[27];PIP3可通过激活Rho 小GTPase家族蛋白调控肌动蛋白骨架系统[28]。因此,推测PTEN 还可能通过改变PIP2和PIP3的浓度及定位,参与紧密连接损伤,但仍需进一步的实验证实。
其次,PTEN作为蛋白质磷酸酶,可以与FAK的第397位酪氨酸位点直接结合,并引起该位点去磷酸化[23]。FAK在睾丸组织内可定位于紧密连接处,与occludin或ZO-1等复合物结合,通过转变其不同位点的磷酸化形式(例如p-FAK-Tyr397、p-FAK-Tyr407、p-FAK-Tyr576)[16-19]参与紧密连接调控。Arp2/3是介导分枝状肌动蛋白细胞骨架成核和聚合的关键蛋白,FAK可以与Arp2/3结合,并增加Arp2/3的活性;而FAK 第397位酪氨酸磷酸化会消减其与Arp2/3之间的相互作用,影响Arp2/3 正常地活化及转移过程,间接造成紧密连接损伤[20,22]。此外,p-FAK-Tyr397水平升高还能进一步促进PI3K/Akt信号通路的激活[29]。因此,由PTEN表达降低引起的p-FAK-Tyr397水平增高及PI3K/Akt信号通路激活并不是两个完全孤立的生物学过程,两者之间密切联系,相辅相成又相互促进,共同参与紧密连接损伤。
综上所述,miR-181d-5p可能通过降低PTEN表达,导致p-FAK-Tyr397水平升高以及激活PI3K/Akt信号通路,参与紧密连接损伤。由于本实验的结果均来自体外实验,存在一定局限性,而紧密连接的动力学调控涉及非常复杂的生物学过程,PTEN 参与紧密连接损伤的确切机制还需要更为详尽的实验证实。
