CNR 转录因子在番茄果实成熟过程中的功能
2021-04-26李玲杨丽霞郭梅
李玲 杨丽霞 郭梅
(天津农学院食品科学与生物工程学院,天津 300384)
成熟果实为人们提供日常生活所需的营养。果实成熟包括一系列的生理代谢过程,在分子层面上果实的生长、发育及成熟是由许多转录因子通过单独或者协同调控特定的下游基因来实现的[1]。SPL(SQUAMOSA promoter-binding protein-like)是植物中特有的转录因子,参与植物叶、花和果实发育,发育时期转化,植物激素和光信号转导,花青素合成,逆境胁迫等过程[2-12]。SPL7 和SPL14 分别通过配置铜元素和控制病原抗性来调节植物的胁迫应答[13]。SPL 调控植物孢子产生和雌雄配体发育[14]。大多数SPL基因具有miR156/157 结合位点,在转录后水平受其调控[15-17]。miR156 调控拟南芥SPL3基因进而在翻译后抑制SPL3 蛋白[17]。miR156/157 的靶基因SPL在番茄茎尖、花和果实中高效表达[18]。Wang等[19]发现SPL 通过MADS box基因调控植物开花时间,提高SPL基因表达水平有助于MADS-box基因家族的转录,促进开花。
CNR 是SPL 转录因子家族中一个比较特殊的成员,已经被证明在果实的发育和成熟过程中起很重要的作用。Manning 等[20]通过使用定点克隆的方法,在第2 条染色体长臂上发现LeSPL-CNR基因。LeSPL-CNR基因启动子区域发生表观遗传变异会影响果皮类胡萝卜素生物合成和细胞壁结构,导致果实不能正常成熟[20]。番茄突变体Cnr(Colourless non-ripening)果实表现出不成熟性状,果皮为黄色,果实类胡萝卜素合成量减少,果实不会变软,与成熟相关酶基因的表达也受到影响[21-22]。Cnr突变体的多聚半乳糖醛酸酶和果胶酯酶的含量及活性比同时期野生型果实低[23]。Karlova 等[24]发现SlAP2a位于LeNAC-NOR、LeMADS-RIN和LeSPL-CNR的下游,LeSPL-CNR 能够与SlAP2a的启动子直接结合,SlAP2a也可以对LeSPL-CNR表达进行反馈调节。总之,CNR 位于RIN、LeHB1、SlAP2a 和 SlTAGL1 的下游,对果实成熟起正调控作用[25]。
目前,由转录因子组成的调控网络是研究果实成熟分子机制的热点,CNR 是调控番茄成熟的关键转录因子。我们团队已经证明CNR 蛋白在破色期番茄果实中表达量最高,发挥了至关重要的作用[26],但是其调控番茄果实成熟的机制和路径尚不清楚。所以,本研究以野生型AC 和突变体Cnr番茄果实为材料,通过转录组测序(RNA-seq)筛选出差异表达基因,然后应用qRT-PCR 技术进一步确定差异基因,最后采用染色质免疫共沉淀技术(ChIP)确定转录因子CNR 直接调控的靶基因,以期为构建以CNR 为中心的果实成熟调控网络奠定理论基础。这不仅能够完善果实成熟的调控理论,而且可以从生物工程角度有效延缓果实成熟,保持果实品质和延长果实贮藏期。
1 材料与方法
1.1 材料
选取由美国康奈尔大学BTI 植物研究所提供的Alisa Craig野生型和Cnr突变体番茄种子,由本实验室繁育保存。番茄种子种植于天津农学院大棚,进行常规管理。选取无机械损害、无病虫害且大小均一的番茄果实进行实验。
1.2 方法
1.2.1 番茄果实总RNA 的提取 采用TRIzol 法提取破色期野生型AC 和突变体Cnr番茄果实总RNA,具体操作步骤依据《分子克隆实验指南》[27]。将组织置于液氮中迅速研磨成干粉,放在10 mL 离心管中,加入4 mL TRIzol 提取液及80 μL β-巯基乙醇,充分混合均匀,室温静置10 min。向提取体系中加入60%氯仿/异戊醇(24∶1),15%无水乙醇,室温静置2 min,4℃ 5 000×g离心15 min。取上清液加入等体积氯仿/异戊醇,4℃ 5 000×g离心10 min,再吸取上清液加入两倍体积的无水乙醇和五分之一体积的3 mol/L NaAC-HAC(pH 5.0)溶液,混合均匀,-20℃静置1h。4℃ 5 000×g离心15 min 得到上清液,加等体积75%乙醇离心5 min,弃去上清,晾干内壁残留的乙醇溶液,用适量DEPC 水溶解沉淀。
1.2.2 RNA-seq 测序及数据分析 从野生型AC 和突变体Cnr番茄果实中提取的RNA 检测合格后,用于构建测序文库,然后送至诺禾致源基因研究中心进行高通量RNA-seq。每个样本3 次生物学重复。最终的数据处理和分析参考Bemer 等[28]的方法。
1.2.3 GO 和KEGG 富集分析 Gene Ontology(简称GO,http://www.geneontology.org)是基因功能国际标准分类体系,由基因本体联合会(Gene Onotology Consortium)建立数据库。基因可以通过ID 对应或者序列注释的方法找到与之对应的GO 编号,而GO 编号可对应到Term,即功能类别或者细胞定位。采用Goseq 软件进行GO 富集分析。KEGG(Kyoto Encyclopedia of Genes and Genomes)是系统分析基因功能、基因组信息,与Pathway 相关的主要公共数据库,是进行生物体内代谢分析、代谢网络研究的强有力工具。应用KOBAS(2.0)软件进行Pathway富集分析。
1.2.4 qRT-PCR 表达差异分析 参照Brown 等[29]的方法进行实时定量PCR 分析,检测差异基因的表达量。qRT-PCR 使用的引物序列见表1。以Actin基因作为内参,每个基因在AC 番茄果实中的表达量归为1。Expression(2-ΔΔCt)表示为平均值±SE。
1.2.5 染色质免疫共沉淀(ChIP)分析 按照Upstate 试剂盒说明书和Li 的方法进行ChIP 分析[30-31]。参照Zhu 等[32]的方法结合GE Health Ni Sepharose six 快速流动亲和层析柱操作说明书进行CNR 蛋白的原核表达和纯化。然后将制备的高纯度、有活性的CNR 蛋白免疫小鼠得到抗CNR 蛋白的多克隆抗体[26]。抗体纯化后用于ChIP 分析。
2 结果
2.1 转录组测序数据的质量评估
野生型AC 和突变体Cnr番茄果实的总RNA 用于构建测序文库,然后在Illumina HiSeq 平台上进行高通量测序。从图1 可以看出,野生型AC 番茄果实3 个重复样本的R2均大于0.974,Cnr突变体番茄果实3 个重复的R2都大于0.971。说明样品的重复性良好,数据非常可靠。
原始图像数据文件通过CASAVA 碱基识别分析转换成Raw Reads,Raw Reads 包含低质量、带接头的Reads。所以Raw Reads 必需经过滤得到Clean Reads 以保证转录组测序数据分析的质量。后续的生物信息学分析都是基于Clean Reads。测序数据质量情况如表2 所示,6 个样本Q20 和Q30 的最小值分别是94.18%和88.92%,表明所有的测序数据均符合生物信息学分析的要求。
选用HISAT 软件将经过拼接的序列进行比较分析和基因组定位。软件的各项参数选择默认值,比对统计结果如表3 所示。各个样本reads mapped 的比例为92.92%-94.77%均高于70%,这说明参考基因组选择恰当,且不存在实验污染的情况,数据和测序结果可信。
2.2 野生型AC和突变体Cnr番茄果实差异基因的筛选结果
图2 是野生型AC 和突变体Cnr番茄果实表达基因的venn 图,以fpkm>1 作为判断基因表达的标准。从图2 中可以看出,AC 表达了17 225 个基因,Cnr表达了19 389 个基因。二者之间共有的表达基因是16 653 个。不同番茄果实差异基因的筛选条件为padj<0.05 且|log2fold change|>1。如图3 所示,野生型AC 和突变体Cnr番茄果实的差异基因有10 223 个,其中上调的差异基因4 341 个,下调的差异基因5 882 个。
从图4 可以看出,与突变体Cnr番茄果实相比,野生型AC 番茄果实中与能量代谢相关具有显著性表达差异的基因是68 个,上调基因28 个,下调基因40 个;有4 个与固碳作用相关的上调差异基因;与线粒体电子传递或ATP 合成相关的基因是11 个差异上调基因,14 个差异下调基因;与脂质代谢有关的基因是22 个差异上调基因和33 个差异下调基因;6 个基因负调控肌醇磷酸盐代谢;与氨基酸代谢相关的是24 个差异上调基因和39 个差异下调基因;存在9 个与核酸代谢有关的差异上调基因和14个差异下调基因;6 个上调基因和6 个下调基因与氧化还原状态有关;2 个下调基因与谷胱甘肽代谢相关;与次生代谢相关的基因有7 个差异上调和10个差异下调;与细胞壁及细胞膜完整性相关的基因包括18 个上调基因和19 个下调基因;3 个基因与番茄红素合成相关,其中2 个上调,1 个下调;4 个上调基因与乙烯生物合成相关;与乙烯信号转导相关的差异基因有4 个上调表达,4 个下调表达;8 个上调差异基因和15 个下调差异基因与植物激素相关;与植物MAPK 信号途径相关的是1 个上调表达的差异基因;3 个下调基因与植物防御应答相关;2个上调和8 个下调基因与植物病原途径相关;3 个上调基因和1 个下调基因与植物非生物胁迫有关;2个差异上调基因与维生素代谢相关。
2.3 差异表达基因GO和KEGG分析
GO 分 为 分 子 功 能(Molecular Function)、生物进程(Biological Process)和细胞组分(Cellular Component)3 部分。通过对野生型和突变体番茄差异表达基因(DEGs)进行GO 富集分析,以揭示哪些生物进程、分子功能和细胞组分与CNR 转录因子相关。如图5 所示,生物进程极显著的集中在次级代谢物生物合成、次级代谢、含硫化合物代谢、α-氨基酸代谢、对高光强度的应答、有机酸生物合成、羧酸生物合成、谷胱甘肽代谢和对脂类的细胞应答;细胞组分显著性的存在于色素体、叶绿体、细胞器亚单位、质体类囊体、质体基质、叶绿体类囊体、类囊体、叶绿体基质、质体包膜、类囊体膜、叶绿体被膜、色素体类囊体膜、叶绿体类囊体膜;分子功能显著性的集中于铁离子束、亚铁血红素结合物、单加氧酶活性、氧化还原酶活性、蛋白丝氨酸/苏氨酸激酶活性、四吡咯结合物、谷胱甘肽转移酶活性、转移酶活性。

表1 用于qRT-PCR 验证的引物序列

图1 番茄果实样本的相关性

表2 数据产出质量情况一览表

表3 Reads 与参考序列比对情况一览表
在生物体内,不同基因相互协调行使其生物学功能,通过Pathway 显著性富集能确定差异表达基因参与的最主要代谢途径和信号转导途径。对差异表达基因进行KEGG Pathway 富集分析,根据P<0.05,选取26 个差异最显著的 KEGG 通路,如图6 所示。野生型AC 和突变体Cnr番茄的差异基因主要富集在谷胱甘肽代谢、MAPK 信号途径、类黄酮化合物代谢、半胱氨酸和甲硫氨酸代谢、类胡萝卜素合成、脂代谢、糖代谢、苯丙素生物合成、萜类化合物合成、植物激素信号转导等途径。

图2 野生型AC 和突变体Cnr 番茄果实差异基因维恩图
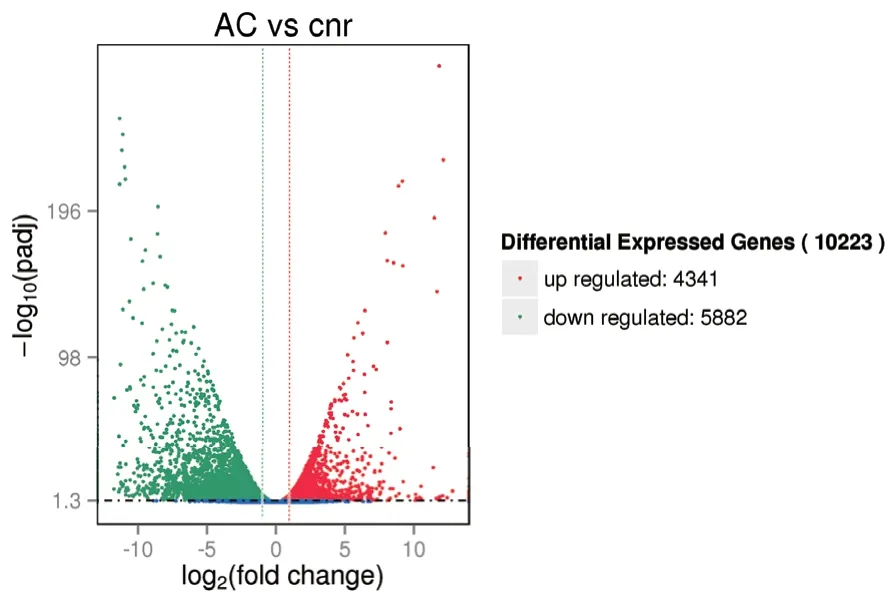
图3 野生型AC 和突变体Cnr 番茄果实基因差异表达分析火山图
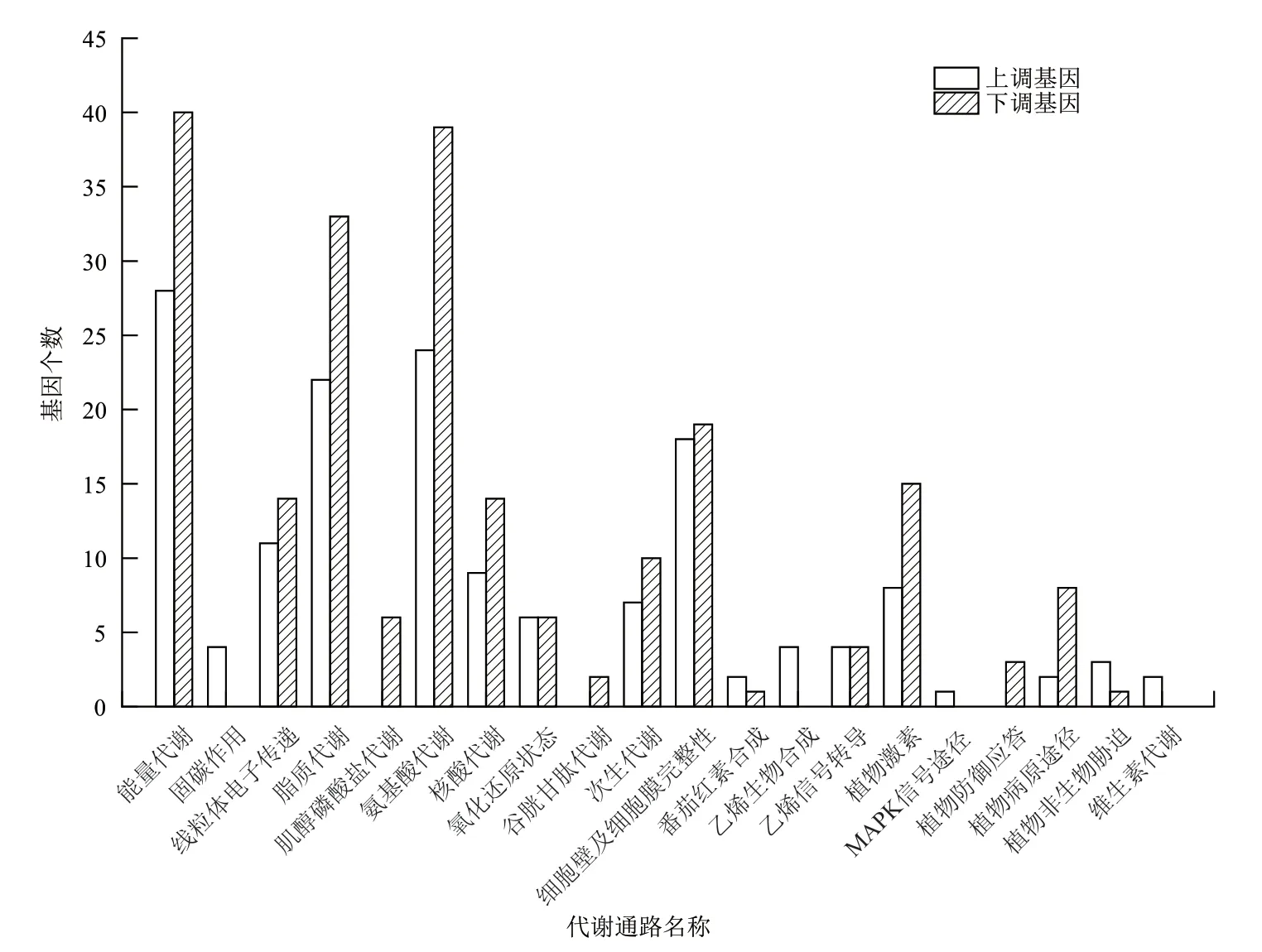
图4 野生型AC 和突变体Cnr 番茄果实差异基因的筛选结果
2.4 qRT-PCR验证AC和Cnr番茄果实中差异基因的相对表达量
选择AC 和Cnr番茄果实中有极显著差异的一些代表性基因进行qRT-PCR 验证。由表4 可知,AC 番茄果实中参与果糖和甘露糖代谢的甘露聚糖-1,4-β-甘露糖苷酶基因(Solyc01g008710.2)的表达量是Cnr番茄果实的2 500 倍。亚油酸9S-脂氧合酶参与亚油酸代谢,AC 番茄果实中该酶基因(Solyc08g014000.2)的相对表达量是Cnr番茄果实的10 000 倍。参与细胞壁果胶代谢的果胶酯酶基因(Solyc07g064180.2)在AC 番茄果实中的相对表达量与亚油酸9S -脂氧合酶基因相似。发现一个新基因(novel.9194)参与番茄红素合成,在AC 番茄果实中的表达量是Cnr番茄果实的100 倍。1-氨基环丙烷-1-羧酸合成酶基因(Solyc05g050010.2)在乙烯生物合成中起重要作用,该基因在AC 番茄果实的表达量是Cnr番茄果实的180 倍。脱落酸和环境应激诱导蛋白TAS14 基因(Solyc02g084850.2)参与植物激素代谢。硫胺素噻唑合酶基因(Solyc07g064160.2)参与硫胺素代谢。促分裂原活化蛋白激酶基因(Solyc02g084870.2)在Cnr 番茄果实中的相对表达量是0.069 7。以上这些基因在AC 番茄果实中上调表达。

图5 野生型AC 和突变体Cnr 番茄果实GO 显著性富集分析

图6 KEGG 显著富集通路
已糖载体基因(Solyc02g079220.2)在Cnr突变体中的相对表达量是101.640 3,钙转运ATP 合成酶基因(Solyc01g096190.2)在Cnr突变体中的相对表达量是6.597 4。脂氧合酶基因(Solyc08g029000.2)在Cnr番茄果实中的表达量是AC 果实中的276 367倍。过氧化物酶基因(Solyc02g082090.2)与氧化还原状态有关,在Cnr番茄果实中的表达量是AC 果实中的1 233 倍。谷胱甘肽S-转移酶基因(Solyc09g011630.2)参与谷胱甘肽代谢,在Cnr番茄果实中的相对表达量是5 383.808 2。果胶酯酶基因(Solyc09g075330.2)在Cnr番茄果实中的表达量是AC 番茄果实中的1 026 倍。番茄红素ε-环化酶基因(Solyc12g008980.1)参与番茄红素代谢,在Cnr突变体中的表达量是15.691 3。乙烯响应蛋白抑制因子1 基因(Solyc08g080630.2)位于乙烯信号转导途径,在Cnr番茄果实中的相对积累量为222.453 4。植物生长素载体组分基因(Solyc07g006900.1)在植物激素代谢过程有重要作用,其在Cnr番茄果实中的相对表达量是 254.237 6,相对表达量较高。葡聚糖-1,3-β-葡萄糖苷酶B 基因(Solyc01g060020.2)与防御应答相关,在Cnr突变体果实中的表达量是AC 果实中的71 倍。
2.5 CNR靶基因启动子区域的ChIP检测
为了证明CNR 在番茄果实体内直接结合的靶基因,我们进行了ChIP 检测。染色质从粉红期果实中提取并与蛋白质交联。用抗CNR 的鼠IgG 免疫沉淀CNR 及其相关DNA-蛋白复合物(ChIP)。阴性对照为正常小鼠的免疫球蛋白G(IgG)。ChIP-qPCR 显示八氢番茄红素合成酶基因(PSY)、多聚半乳糖醛酸酶基因(PG)、1-氨基环丙烷-1-羧酸合成酶基因(ACS)、亚油酸9S-脂氧合酶基因和果胶酯酶基因的GTAC 基序显著富集(图7)。八氢番茄红素合成酶基因的结合能力最强,果胶酯酶基因结合能力最弱。ChIP-qPCR 结果说明在番茄果实体内CNR 直接结合在PSY、ACS、LOX、PE以及PG基因的启动子区域,这些基因是转录因子CNR 的靶基因。
3 讨论
番茄果实的成熟和衰老过程是复杂而有序的,包括颜色、质地、风味、香气成分和其他新陈代谢的变化,同时又与基因调控过程高度相关。野生型AC 和突变体Cnr番茄果实的差异基因涉及番茄果实生长发育的多个方面,包括能量相关代谢途径、固碳作用、线粒体电子传递、ATP 合成、脂代谢、磷酸肌醇代谢、氨基酸代谢、核酸代谢、氧化还原状态、谷胱甘肽代谢、次生代谢、细胞壁、细胞膜完整性、番茄红素合成、乙烯生物合成、乙烯信号转导、植物激素、植物防御应答、植物病原相关、非生物胁迫、维生素代谢和MAPK 信号通路。
脂氧合酶(LOX)是高等植物中的一种非血红素铁蛋白,参与香气成分合成、果实成熟和衰老、氧胁迫、脂质过氧化等过程,对细菌感染和不利的外部条件有很好的防御作用[33-34]。LOX 的反应产物可能通过参与乙烯对跃变型果实的触发而加速果实成熟。另一方面,LOX 通过催化不饱和脂肪酸的氧化影响植物成熟和衰老。亚油酸9S-脂氧合酶基因(Solyc08g014000.2)在AC 番茄果实中表达量较高,脂氧合酶基因(Solyc08g029000.2)的相对表达量在AC 番茄果实中降低。果胶酯酶在植物组织和微生物中分布较为广泛,作用于细胞壁降解、细胞分裂,果实软化及植物抗病[35]。与Cnr突变体番茄果实相比,AC 番茄果实中的果胶酯酶基因(Solyc07g064180.2)上调,另一个果胶酶基因(Solyc09g075330.2)下调。以上结果表明,脂氧合酶和果胶酯酶是由多基因家族编码的,各个基因在番茄果实成熟和软化过程中起不同作用。

表4 AC 和Cnr 番茄果实差异基因的相对表达量

图7 鉴定CNR 在番茄果实体内结合的顺式元件
过氧化物酶催化涉及过氧化氢的各种氧化反应,有多重氧化还原进程。谷胱甘肽过氧化物酶基因(Solyc08g080940.2)参与上述反应。过氧化物酶的积累可以应对由不良伤害和非生物胁迫引起的活性氧(ROS)过量产生。过氧化物酶基因(Solyc02g082090.2)在AC 番茄果实中下调表达。谷胱甘肽转移酶(GST)属于蛋白质超家族,当植物受到非生物胁迫时,它可以清除活性氧并保护植物细胞膜结构完整和蛋白质的活性[36]。AC 番茄果实的谷胱甘肽 S-转移酶基因(Solyc09g011630.2)下调。丝裂原激活蛋白激酶(MAPK)信号通路在植物免疫反应中起重要作用,该酶基因(Solyc02g084870.2)在Cnr突变体番茄果实中没有表达,仅在AC 番茄果实中检测到,这揭示CNR 转录因子与MAPK 途径密切相关。番茄红素具有抗氧化性能,能够保护机体免受氧化带来的损伤,同时可以介导细胞通讯对内分泌和免疫系统进行调控[37]。本研究发现了一个与番茄红素合成相关的新基因(novel.9194),该基因在AC 番茄果实中上调表达;番茄红素ε-环化酶基因(Solyc12g008980.1)在AC 番茄果实中下调表达,这两个基因在番茄果实的着色过程中共同起作用。
乙烯是一种激素,影响成熟相关基因的表达,在水果的生长过程中起重要的调节作用[38]。AC 番茄果实中ACS基因(Solyc05g050010.2)呈上调表达,Cnr突变体中该基因不表达,无乙烯合成。乙烯响应蛋白抑制因子1 基因(Solyc08g080630.2)在AC番茄果实中呈负调控。LOX 途径是植物特有的芳香物质合成途径,一些挥发性化合物通过LOX 途径合成,如LOX 通过亚麻酸途径参与醛香物质的合成[39]。也有研究发现,在香气物质的形成过程中乙烯发挥了重要作用。LOX 途径中与香气相关的基因Tomlox C表达降低,说明番茄果实LOX 途径中香气物质的产生依赖乙烯[40]。氨基酸代谢途径的差异表达基因决定野生型AC 和突变体Cnr番茄果实具有不同风味,但是番茄果实香气的组成和形成机理仍需进一步研究。
4 结论
CNR 转录因子的功能涉及番茄果实生长发育阶段的各个方面。CNR 通过直接结合八氢番茄红素合成酶基因(PSY)、1-氨基环丙烷-1-羧酸合成酶基因(ACS)、亚油酸9S-脂氧合酶基因(LOX)、果胶酯酶基因(PE)以及聚半乳糖醛酸酶基因(PG)的启动子来调控番茄的成熟软化过程。