外引大麦农艺性状SSR关联位点及等位变异表型效应分析
2018-05-11吕仲昱贾建磊卢九斤王晋民朱惠琴杨莉娜李宗仁
赖 勇,吕仲昱,贾建磊,卢九斤,王晋民,朱惠琴,马 辉,田 丰,杨莉娜,李宗仁
(1.青海大学农牧学院,青海西宁 810016;2.三江源生态与高原农牧业国家重点实验室,青海西宁 810016;3.甘肃隆源农业科学研究所,甘肃兰州 730070)
大麦是青藏高原的主栽作物之一。遗传基础狭窄是目前大麦育种家面临的严重困境[1],而引进种质资源是解决该问题的有效手段。借助分子标记技术对引进的种质资源进行遗传分析,发掘不同材料所含的有利等位变异,对于作物育种的亲本选择、组合配制和标记辅助育种具有重要意义。基于SSR标记的遗传多样性、基因定位和关联分析在各类作物中广泛应用[2-4]。国内外也有许多关于大麦SSR遗传多样性分析的报道[5-7]。通过关联分析寻找与目标性状相关的位点,分析其对目标性状的贡献率是发掘有利基因的主要方法[8-12]。但是目前,大量关联分析都停留在标记或位点水平,鲜有具体分析相关位点各等位变异的表型效应。对于SSR位点,在自然群体中通常能检测到多个等位变异,每个等位变异的遗传效应存在差异[13-14]。分析每一个等位变异的表型效应对于分子标记辅助选择育种具有指导意义。Breseghello和Sorrells[13]在进行小麦籽粒直径及磨粉品质与SSR标记关联分析中,以无效等位基因(null allele)材料为对照,分析各等位变异的表现效应值,发掘出其中可以增大籽粒直径和提高磨粉品质的有利基因。此方法给作物关联作图中,具体分析不同等位基因的表型效应、发掘有利基因提供了新思路。Zhang等[14]利用Breseghello和Sorrells[13]的方法寻找到降低小麦株高的有利等位变异。文自翔等[15]采用该方法从我国栽培大豆和野生大豆中鉴定出许多与农艺性状以及品质性状相关联的优异等位变异。在大麦上,还未见基于SSR位点对引进种质不同等位变异表型效应进行分析的报道。
本研究在以前对大麦引进国外种质资源进行遗传多样性和GLM关联分析[16]的基础上,进一步采用MLM关联分析,检测大麦农艺性状关联位点,并分析关联位点上不同等位变异的表型效应,以期为青海省大麦分子标记辅助选择提供一定的参考。
1 材料与方法
1.1 种植材料及农艺性状鉴定
55份国外引进大麦品种包括19份丹麦品种、12份匈牙利品种、12份德国品种以及少量其他国外引进栽培品种,分别于2014年3月29日、2015年3月28日种植于青海大学农牧学院试验地。这些材料为没有直接亲缘关系的自然群体,符合关联分析发掘有利等位变异的条件。材料名称、来源及种植情况等详见文献[16]。于分蘖期采幼嫩叶片,-80 ℃保存备用。在各材料成熟后,每个材料随机抽取10个单株,测定株高、穗长、穗下茎长、千粒重、全生育期等农艺性状。各材料具体农艺性状指标已在文献[16]中报道。
1.2 混合模型关联分析
通过软件SPAGeDi构建亲缘关系矩阵(K),结合前文GLM分析中获得的Q参数[16],以Q+K的方法采用混合线性模型(mixed linear model,MLM)在Tassel 2.1 软件中运行,检测与农艺性状相关联的标记,P<0.01时为阳性关联位点。以软件Origin 8.0绘制关联位点图。
1.3 等位变异表现效应分析
参考Breseghello和Sorrells[13]提出的无效等位变异(null allele)方法,区分有效等位变异和无效等位变异,对MLM模型中与农艺性状相关联的位点,分析其不同等位变异的表型效应。具体表型效应计算方法:
ai=∑xij/ni-∑Nk/nk
式中ai为第i个等位变异的表型效应值,xij表示含有第i个等位变异的第j个材料性状表型测定值,ni为含有第i个等位变异的材料数,Nk为携带无效等位变异的第k个材料的表型测定值,nk为具有无效等位变异的材料数。若ai值为正, 则认为该等位变异为正向(或增效)等位变异,反之为负向(或减效)等位变异。以软件Origin 8.0绘制效应值柱形图。
当某性状同时有多个正向(或负向)等位变异或多个相关联位点时,参考文自翔等[15]方法计算各位点全部增效或减效等位变异的平均效应[average positive(negative)allele effect of locus,AAE],评价该位点的整体增效或减效能力。
AAE=∑ac/nc
式中ac为某关联位点内第c个增效(或减效)等位变异表型效应值,nc为位点内增效(或减效)等位变异数。增效(或减效)比例:AN=AAE∑NK/nK×100%
2 结果与分析
2.1 MLM关联分析结果
经MLM关联分析,共检测到15个与株高、穗长、穗下茎长、千粒重和全生育期关联的位点(P<0.01),其中位点GBMS2和HVM33均与穗长和千粒重相关联,位点HVM33、HVM60和Bmac316均与千粒重相关联(P<0.001)(图1、表1)。各位点分别位于除1H、5H外的其余5条染色体上,不同位点对表型变异的解释率变化范围为3.19%~24.30%(表1)。与千粒重相关联的位点最多,有6个,只有一个位点HV13GEIII与全生育期关联。

图1 与所测农艺性状相关联的15个SSR位点
Table1ContributionoflocustoagronomictraitsunderMLM
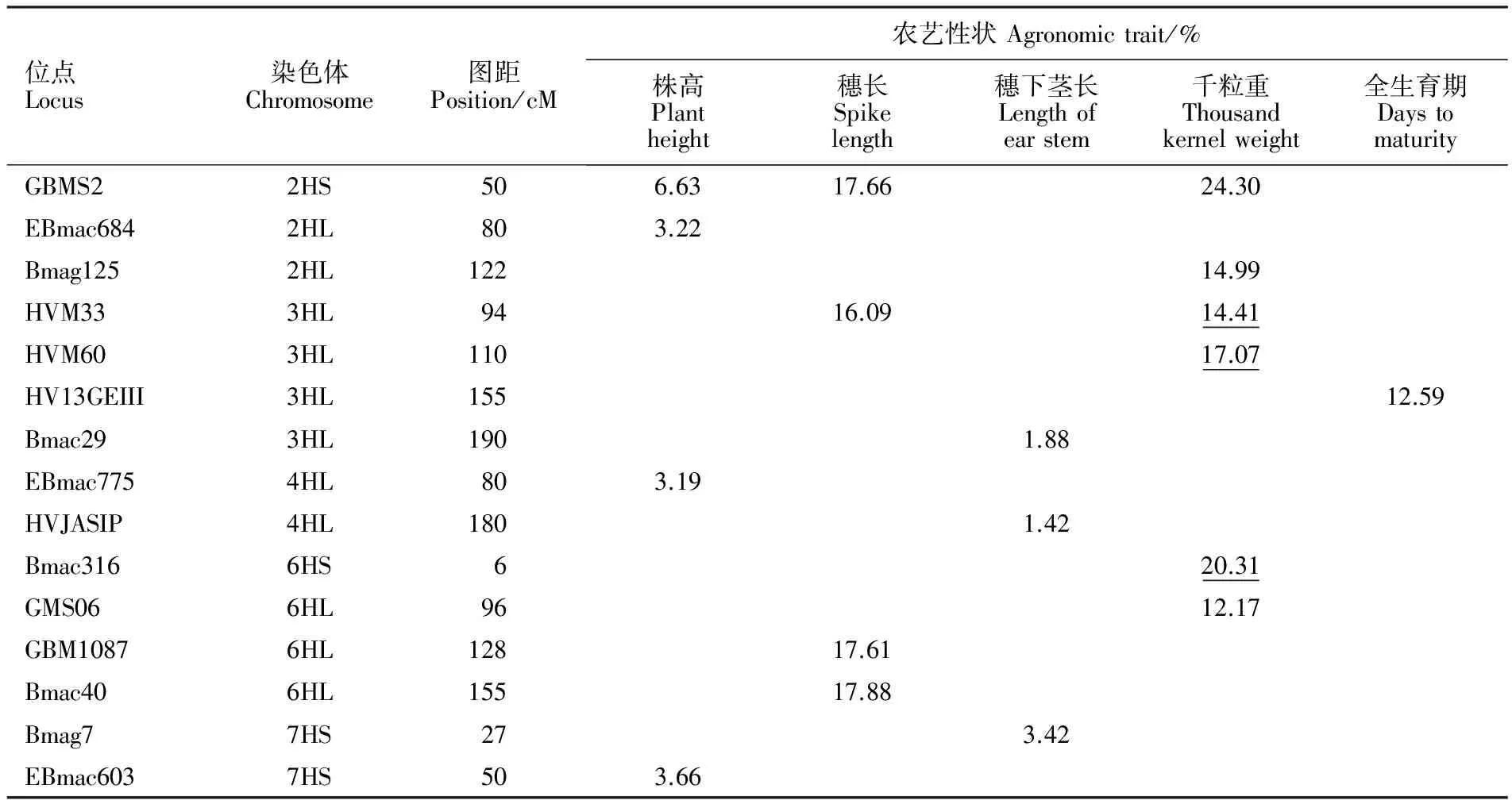
位点Locus染色体Chromosome图距Position/cM农艺性状Agronomictrait/%株高Plantheight穗长Spikelength穗下茎长Lengthofearstem千粒重Thousandkernelweight全生育期DaystomaturityGBMS22HS506.6317.6624.30EBmac6842HL803.22Bmag1252HL12214.99HVM333HL9416.0914.41HVM603HL11017.07HV13GEIII3HL15512.59Bmac293HL1901.88EBmac7754HL803.19HVJASIP4HL1801.42Bmac3166HS620.31GMS066HL9612.17GBM10876HL12817.61Bmac406HL15517.88Bmag77HS273.42EBmac6037HS503.66
下划线表示在0.001水平相关联位点。
The underlined number showed locus associated with agronomic trait at 0.001 level.
2.2 等位变异表型效应分析
15个关联位点中,HVM33、Bmac40、EBmac775、EBmac603、Bmag125、HVM60和Bmac1367位点上检测到无效等位基因。这7个位点上共有22个不同有效等位变异,其中Bmac40-2、EBmac755-2和EBmac755-4等6个等位变异对表型具有正向效应,其余均为负向效应(图1)。与穗长相关联的位点HVM33和Bmac40上共检测到1个具有正向效应的等位变异和6个具有负向效应的等位变异(图2A),且负向效应明显大于正向效应。与株高相关联的位点上共检测到3个具有正向效应的等位变异和5个具有负向效应的等位变异(图2B),其中等位变异EBmac775-4的正向效应值最大(+31.7 cm),EBmac603-5具有最大负向效应(-25.6)。与千粒重相关联的3个位点上共检测到10个有效等位变异,其中2个为正向效应,8个为负向效应(图2C)。
2.3 整体平均效应分析结果
在7个相关联位点中,等位变异平均效应以负向为主(表2)。与穗长相关联的2个位点HVM33和Bmac40上,等位变异平均效应均为负向,且平均效应差异较小,分别为-1.6 cm 和-1.0 cm。与株高相关联的位点中,EBmac775的等位变异平均效应为+16.91 cm,EBmac603为-13.35 cm。与千粒重相关联的位点中,HVM60等位变异平均效应为正向,HVM33、Bmag125和Bmac316三位点的等位变异平均效应均为负向。

图2 穗长、株高和千粒重关联位点等位变异表型效应
Table2Averageeffectsofallelicvariationsassociatedwithspikelength,plantheightandthousandkernelweight
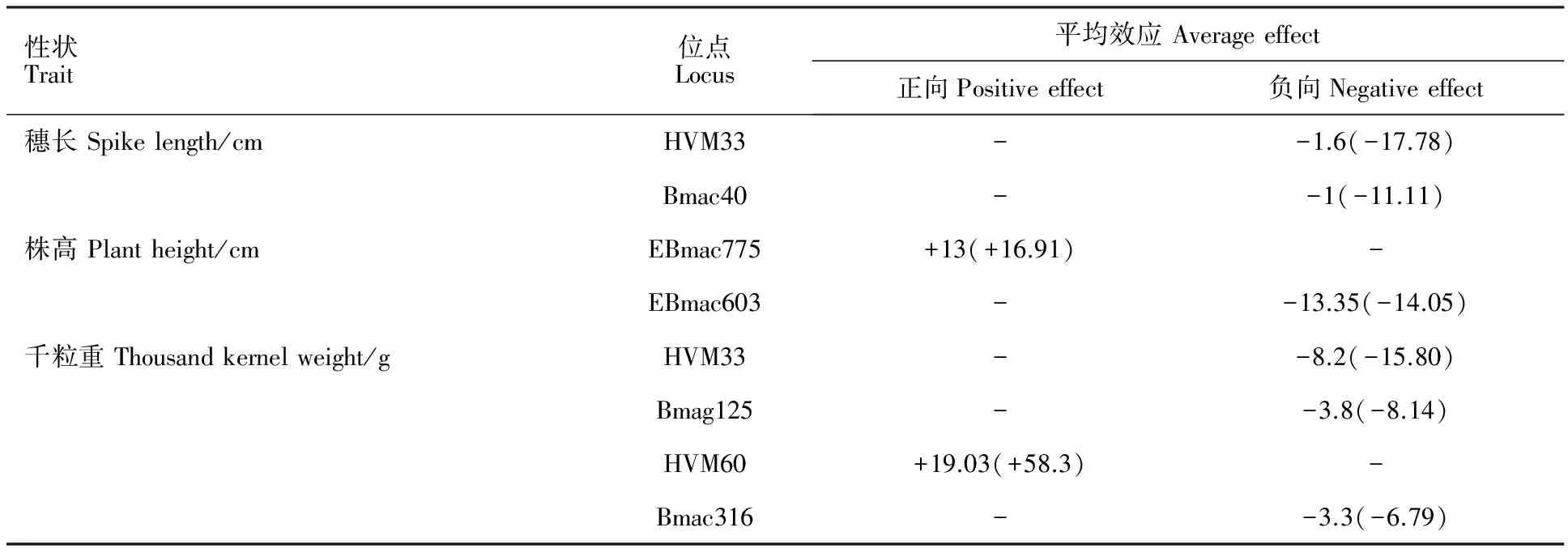
性状Trait位点Locus平均效应Averageeffect正向Positiveeffect负向Negativeeffect穗长Spikelength/cmHVM33--1.6(-17.78)Bmac40--1(-11.11)株高Plantheight/cmEBmac775+13(+16.91)-EBmac603--13.35(-14.05)千粒重Thousandkernelweight/gHVM33--8.2(-15.80)Bmag125--3.8(-8.14)HVM60+19.03(+58.3)-Bmac316--3.3(-6.79)
括号内为与无效等位变异相比较的平均正向(或负向)效应占的比例(%)。
The number in parentheses indicates the proportion(%) of average positive(negative) allele effect of locus compared with the effect of null allele.
3 讨 论
对引进种质资源进行关联分析以发掘有利等位变异,是进行分子标记辅助选择的基础。本研究采用SSR标记技术对国外引进的大麦品种进行MLM关联分析和等位变异表型效应分析,为后期大麦分子标记辅助育种提供了一定的理论依据。一些研究结果表明,关联分析中的MLM模型比GLM模型的结果准确,可以更有效地降低伪关联[17-19]。本研究在前文的基础上进一步采用MLM模型进行关联分析,共检测到15个与株高、穗长、千粒重等农艺性状相关联的位点,与GLM模型相比[16],检测到的关联位点有所减少,且各位点对表型的解释率明显低于GLM模型。因此,在进行等位变异的表型效应分析中,集中于MLM模型中检测到的关联位点。
具体分析某一位点等位变异的效应值,有助于发掘其中的有利等位变异,为分子标记辅助育种提高更详细的依据。魏添梅等[20]以抗旱小麦材料为对象进行分子标记与株高的关联作图时,以优良材料含有的等位变异作为优异等位变异。但是这种方法不能具体分析每一个等位变异的表型效应值,不能给分子标记后期辅助选择提供更翔实的依据。这种分析方法仅对于没有检测到无效等位变异的位点具有一定的借鉴意义。当检测到无效等位变异的位点时,采用Breseghello和Sorrells[13]的方法更具优势。因此在进行等位变异表型效应分析时,可以综合两种方法评价种质资源所含等位变异的优异程度。本研究从不同位点上共检测到7个无效等位变异,以之为对照,对每个有效等位变异的表型效应进行具体分析。穗长关联位点上,只有等位变异Bmac40-2具有正向效应(+0.4 cm),是增加穗长优异等位变异。株高关联的位点上,有5个可以降低株高的优异等位变异,以等位变异EBmac603-4(-17.6 cm)和EBmac603-5(-25.6 cm)最显著。千粒重关联的等位变异中,只有HVM60-1和HVM60-12,为正向效应,效应值分别为+13.5 g和+12.0 g,可以作为提高千粒重的优异候选等位变异。在分子标记后期标记辅助选择时,在穗长方面,可以标记Bmac40进行检测,含有Bmac40-2等位变异的个体则具有较大穗长的趋势;在千粒重方面,可以标记HVM60进行检测,含有HVM60-1和HVM60-12等位变异个体的千粒重将更大。
参考文献:
[1]TANKSLEY S D,MCCOUCH S R.Seed banks and molecular maps:unlocking genetic potential from the wild [J].Science,1997,277(5329):1063.
[2] 梁银培,孙 健,索艺宁,等.水稻耐盐性和耐碱性相关性状的QTL定位及环境互作分析[J].中国农业科学,2017,50(10):1747.
LIANG Y P ,SUN J,SUO Y N,etal.QTL mapping and QTL × environment interaction analysis of salt and alkali tolerance-related traits in rice(OryzasativaL.) [J].ScientiaAgriculturaSinica,2017,50(10):1747.
[3] INGHELANDT D V,MELCHINGER A E,LEBRETON C,etal.Population structure and genetic diversity in a commercial maize breeding program assessed with SSR and SNP markers [J].TheoreticalandAppliedGenetics,2010,120(7):1289.
[4] WRSCHUM T,LANGER S M,LONGIN C F,etal.Population structure,genetic diversity and linkage disequilibrium in elite winter wheat assessed with SNP and SSR markers [J].TheoreticalandAppliedGenetics,2013,126(6):1477.
[5] WANG A,YU Z,DING Y.Genetic diversity analysis of wild close relatives of barley from Tibet and the Middle East by ISSR and SSR markers [J].ComptesRendusBiologies,2009,332(4):393.
[6] MALYSHEVA-OTTO L,GANAL M W,LAW J R,etal.Temporal trends of genetic diversity in European barley cultivars(HordeumvulgareL.) [J].MolecularBreeding,2007,20(4):309.
[7] KRAAKMAN A T W,MARTNEZ F,MUSSIRALIEV B,etal.Linkage disequilibrium mapping of morphological,resistance,and other agronomically relevant traits in modern spring barley cultivars [J].MolecularBreeding,2006,17(17):41.
[8] ZONDERVAN K T,CARDON L R.The complex interplay among factors that influence allelic association [J].NatureReviews.DrugDiscovery,2004,5(2):89.
[9] GUPTA P K,RUSTGI S,KULWAL P L.Linkage disequilibrium and association studies in higher plants:Present status and future prospects [J].PlantMolecularBiology,2005,57(4):461.
[10] IVANDIC V,HACKETT C A,NEVO E,etal.Analysis of simple sequence repeats(SSRs) in wild barley from the Fertile Crescent:Associations with ecology,geography and flowering time [J].PlantMolecularBiology,2002,48(5-6):511.
[11] KRAAKMAN A T W,MARTNEZ F,MUSSIRALIEV B,etal.Linkage disequilibrium mapping of morphological,resistance,and other agronomic relevant traits in modern spring barley cultivars [J].MolecularBreeding,2006,17(17):41.
[12] SUN D F,REN W B,SUN G L,etal.Molecular diversity and association mapping of quantitative traits in Tibetan wild and worldwide originated barley(HordeumvulgareL.) germplasm [J].Euphytica,2011,178(1):31.
[13] BRESEGHELLO F,SORRELLS M E.Association mapping of kernel size and milling quality in wheat(Triticumaestivum) cultivars [J].Genetics,2006,172(2):1165.
[14] ZHANG J,HAO C,REN Q,etal.Association mapping of dynamic developmental plant height in common wheat [J].Planta,2011,234(5):891.
[15] 文自翔,赵团结,郑永战,等.中国栽培和野生大豆农艺及品质性状与SSR标记的关联分析II.优异等位变异的发掘[J].作物学报,2008,34(8):1339.
WEN Z X,ZHAO T J,ZHENG Y Z,etal.Association analysis of agronomic and quality traits with SSR markers inGlycinemaxandGlycinesojain China:II.Exploration of elite alleles [J].ActaAgronomicaSinica,2008,4(8):1339.
[16] 赖 勇,贾建磊,王晋民,等.外引大麦SSR标记遗传多样性及其与农艺性状的关联分析[J].麦类作物学,2017,37(2):197.
LAI Y,JIA J L,WANG J M,etal.Analysis of genetic diversity and association with agronomic traits in barley(HordeumvulgareL.) introduced from abroad using SSR markers [J].JournalofTriticeaeCrops,2017,37(2):197.
[17] YANG X H,YAN J B,SHAH T,etal.Genetic analysis and characterization of a new maize association mapping panel for quantitative trait loci dissection [J].TheoreticalandAppliedGenetics,2010,121(3):417.
[18] ZHANG Z W,ERSOZ E,LAI C Q,etal.Mixed linear model approach adapted for genome-wide association studies [J].NatureGenetics,2010,42(4):355.
[19] PRICE A L,ZAITLEN N A,REICH D,etal.New approaches to population stratification in genome-wide association studies [J].NatureReviewsGenetics,2010,11(7):459.
[20] 魏添梅,昌小平,闵东红,等.小麦抗旱品种的遗传多样性分析及株高优异等位变异挖掘[J].作物学报,2010,36(6):895.
WEI T M,CHANG X P,MIN D H,etal.Analysis of genetic diversity and tapping elite alleles for plant height in drought-tolerant wheat varieties [J].ActaAgronomicaSinica,2010,36(6):895.
