胍基乙酸在猪营养中的研究进展
2021-04-12王向明刘则学
王向明,刘则学
[1.中粮家佳康(张北)有限公司,河北 张家口 076450;2.武汉中粮肉食品有限公司,湖北 武汉 430200]
胍基乙酸(guanidine acetic acid, GAA)又名为胍乙酸、N-咪基甘氨酸,是肌酸的天然前体[1],其理化性质见表1。动物机体中,L-精氨酸和甘氨酸可以合成胍基乙酸,L-精氨酸咪基被L-精氨酸甘氨酸咪基转移酶分开,L-精氨酸转变为L-鸟氨酸,甘氨酸与切下的咪基结合形成胍基乙酸[2]。肌酸在肝脏中是由GAA甲基化形成的[3],在饲粮中加入GAA的目的是为了增加动物体内肌酸含量,欧盟在2009年10月将GAA列入饲料添加剂目录[4]。
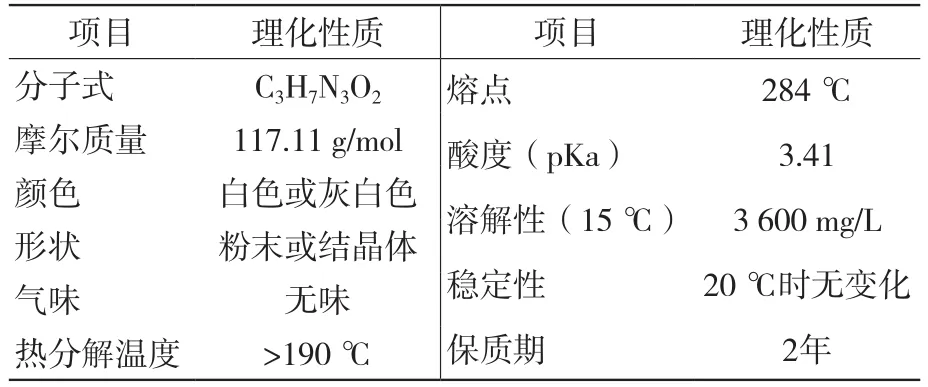
表1 GAA理化性质
1 GAA的代谢途径
胍基乙酸约80年前首次被确定为人类的天然化合物[5]。20世纪50年代被用作治疗剂[6]。此后的研究发现,甘氨酸和L-精氨酸在动物体内会形成GAA,即L-精氨酸甘氨酸咪基转移酶(L-arginine:glycine amidinotranferase, AGAT)将L-精氨酸的咪基分开,形成L-鸟氨酸,被切下的咪基与甘氨酸结合形成GAA,所以GAA也被称作甘氨酸衍生物,存在于动物的各组织器官[7],且还是尿液的常规成分[8]。在人体中,肾脏合成的GAA约占GAA总产量的20%,表明肾脏是合成GAA的主要场所,但GAA也在其他组织(胰腺、肝脏和肌肉)中合成[9]。自身合成或从饲粮中吸收的GAA,都可通过血液循环转移至肝脏,利用甲基化的方式合成肌酸。在肝脏中,GAA在生物转化过程中需要S-腺苷蛋氨酸-胍基乙酸N-甲基转移酶(S-adenosylmethionine:guanidinoacetate N-methyltransferase, GAMT)的催化,S-腺苷蛋氨酸(S-adenosyl methionine, SAM)可给GAA提供甲基,GAA与甲基结合生成肌酸和S-腺苷高半胱氨酸(S-adenosyl-L-homocysteine, SAH),在腺嘌呤核苷同型半胱氨酸水解酶的作用下,SAH可生成腺嘌呤核苷和同型半胱氨酸。GAA合成肌酸所需要的甲基不是由机体内其他甲基化供给,主要是由SAM提供[3],且由SAM提供甲基的反应85%以上在肝脏中进行[10]。据报道,约40%的甲基用于通过GAMT催化的反应合成肌酸[11]。肌酸在体内可转变成磷酸肌酸和肌酐,参与能量代谢。研究发现,L-精氨酸和甘氨酸在合成胍基乙酸的过程中分别作为机体合成肌酸的限速步骤和调节靶点[12]。肌酸可利用反向调节功能调整甘氨酸咪基转移酶的表达[13],但不能反向调节S-腺苷蛋氨酸-胍基乙酸N–甲基转移酶的表达[12]。这表明胍基乙酸和肌酸的合成步骤是不可逆的,必须胍基乙酸合成在前,肌酸合成在后。如果GAA在饲粮中过量添加,就会造成甲基需要量增大,导致同型半胱氨酸量增多,易引发胱氨酸血症[14-15]。
2 GAA的生物学功能
2.1 GAA促进动物代谢合成
GAA能够刺激或抑制与糖代谢有关的关键酶的活性加强体内分解代谢,增加三磷酸腺苷(adenosine triphosphate, ATP)水平从而来加强能量物质的合成[16]。此外,机体合成肌酸的主要基础物质就是GAA,作为体内新陈代谢的重要组成部分,肌酸是能量临时储存的场所。肌酸包括游离肌酸和磷酸肌酸,两者在体内共同组成磷酸原系统。当机体ATP含量高时,磷酸肌酸就会把能量储存;当机体ATP含量低时,磷酸肌酸就会释放ATP,但因ATP无法穿透细胞膜,因此不能从体外获得ATP,所以只能由葡萄糖或其他能源物质提供[17]。合成肌酸的前体物质就是胍基乙酸,生产中可以在饲粮中加入胍基乙酸来达到补充肌酸的目的。在肌酸激酶的作用下,肌酸可以形成磷酸肌酸,磷酸肌酸一方面是机体活细胞能量来源的关键物质,另一方面也是机体组织中关键的能量储备[18-19],机体缺乏能量时,可以利用肌酸激酶催化磷酸肌酸,使其释放ATP,从而提高机体ATP水平[20]。
2.2 GAA在动物抗氧化过程中的作用
利用抗氧化系统,动物能快速消除机体过多的自由基,从而维持机体自由基平衡。正常情况下,自由基的生成和清除是处于平衡状态的。在受到外部刺激或机体紊乱时,自由基会大量产生并积聚,当超出机体抗氧化系统的耐受力后,就会导致机体发生氧化损伤,从而影响动物的健康。饲喂含有GAA饲粮的动物体内抗氧化活性增强有两个原因:一是摄取GAA后,动物体内肌酸和磷酸肌酸含量明显增加,肌酸具备良好的抗氧化功能,磷酸肌酸能够加强抗氧化酶系统功能,以此来防止氧化应激的出现。肌酸可通过清除活性氧,特别是羧基和活性氮来保护细胞,此外,肌酸激活肌酸激酶(creatine kinase, CK)利用二磷酸腺苷(adenosine diphos-phate, ADP)再循环系统消除自由基,从而起到抗氧化功能[21]。Lawler等[22]研究表明,高浓度肌酸能够清除自由基;Guidi等[23]发现,由氧化应激造成的线粒体DNA损伤可通过肌酸得到缓解。二是添加GAA可减少动物的内源性合成,降低精氨酸消耗,增加细胞内精氨酸的含量。精氨酸具有抗氧化作用,可以消除如超氧阴离子等自由基,能够清除黄嘌呤氧化酶所产生的超氧阴离子,抑制铜离子诱发的脂蛋白氧化和内皮细胞及主动脉环释放的超氧阴离子[24]。此外,内皮细胞中氧化低密度脂蛋白(low density lipoprotein, LDL)可诱导机体发生氧化应激,而精氨酸则可阻止机体受到损伤[25]。GAA是合成肌酸的前体物质,在动物饲粮中添加GAA可以提高机体肌酸和磷酸肌酸的量,节省精氨酸的消耗,GAA就是利用这种方式达到抗氧化的目的。
3 GAA在猪饲粮中的应用
3.1 GAA对猪生产性能的影响
饲粮中添加GAA可以有利于肌酸的合成,提高猪的生产性能[26]。相关研究见表2。
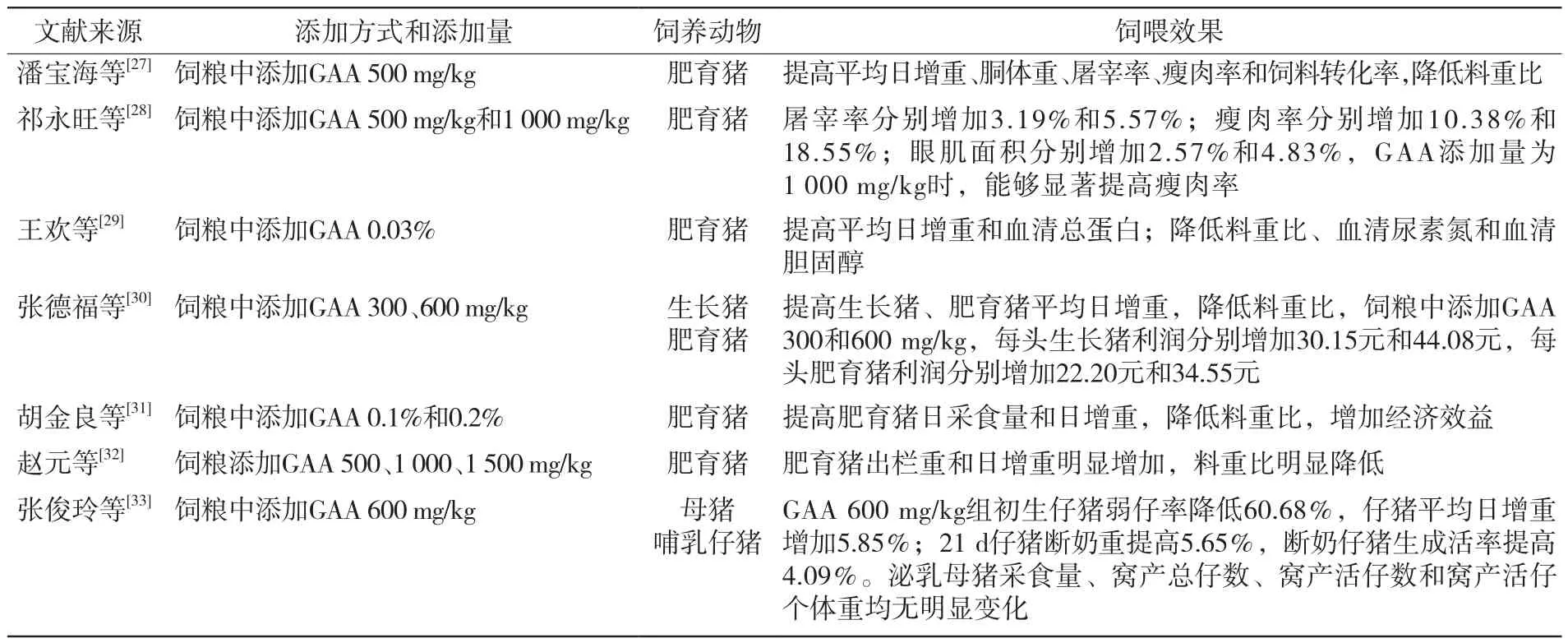
表2 GAA对猪生产性能的影响
饲粮中加入GAA有助于动物生长,原因可能是GAA的内源合成减少了。GAA在体内合成需要甘氨酸和精氨酸的参与,在饲粮中添加GAA可以减少甘氨酸和精氨酸的消耗量,这样使得精氨酸和甘氨酸可以最大程度地参与机体氨基酸和蛋白质的合成,从而促进机体的生长[34]。作为生长肥育猪的必需氨基酸,当饲粮中精氨酸缺乏时可以通过添加GAA来解决。机体摄入GAA后,可以增加肌肉中肌酸和磷酸肌酸的含量,降低机体碳水化合物、脂肪和蛋白质的损耗。GAA一方面影响γ-氨基丁酸分泌,促进下丘脑分泌促生长激素释放激素,进而促进垂体分泌生长激素[1];另一方面,GAA还可以促进类胰岛素样生长因子-Ⅰ(IGF-Ⅰ)的分泌来促进机体生长[35]。
通过表2可知,饲粮中添加GAA能提高仔猪和肥育猪日增重和降低料重比,并且添加GAA可以提高磷酸肌酸和ATP等利用率,减少蛋白质、脂肪和碳水化合物的分解,进而有助于促进动物生长,提高饲料利用率。
3.2 GAA对猪胴体品质和肉质的影响
在动物饲粮中添加GAA,不仅可以提高胴体性状,而且可以改善肉质。有关研究如表3所示。
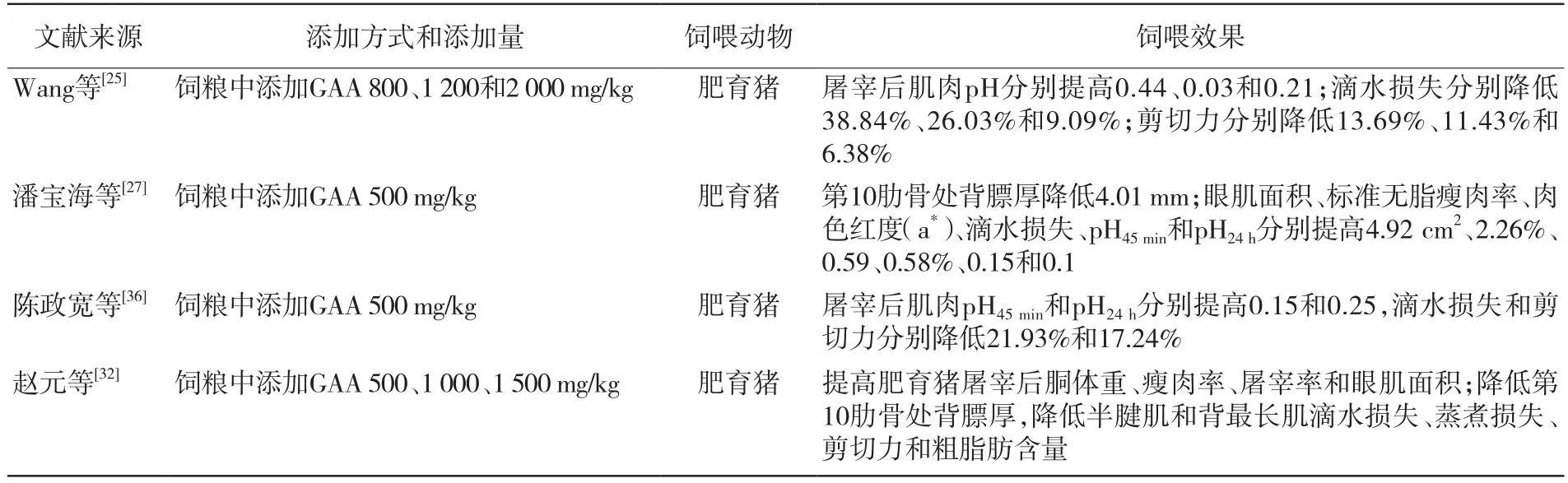
表3 GAA对猪胴体性状和肉质的影响
综上所述,饲粮中添加GAA对肥育猪生长性能、胴体性状、肉质、肌肉品质的改善具有积极作用。通常,动物屠宰后其血液内循环系统就会停止供氧,造成ATP水平下降,此时无氧酵解发挥作用,在肌肉中产生乳酸,导致肌肉pH降低。然而,在饲粮中添加GAA可以增加肌酸和磷酸肌酸的储备,提高ATP的含量,屠宰后可以缓解无氧酵解在肌肉中的反应,进而提高肌肉pH。Rahman等[37]发现,肌肉系水力增加能够改善肉的硬度。屠宰后肌肉pH达到最低前,钙离子会从肌细胞的肌浆内质网中游离出来[38],在能量的作用下会出现肌肉收缩。通常,当肌肉收缩达到40%时,硬度最高,但当收缩大于40%时,肌肉会变得柔软,导致这一结果的原因是肌动蛋白丝发生Z线断裂造成的,添加GAA可以增加ATP的水平,在能量充足的条件下肌肉产生超收缩,进而提升肌肉嫩度[35]。
3.3 GAA对猪抗氧化性能和糖代谢关键酶的影响
动物利用抗氧化系统能够快速清除机体过量的自由基,保持机体稳态。丙二醛(Malondialdehyde,MDA)作为脂质过氧化的终产物可以体现机体过氧化程度。作为体内抗氧化酶之一的超氧化物歧化酶(Superoxide Dismutase,SOD),能够把超氧根阴离子歧化为过氧化氢(Hydrogen peroxide, H2O2),从而清除毒性[39]。谷胱甘肽(Glutathione, GSH)能够消除体内自由基,总抗氧化能力(Total Antioxidant Capacity, T-AOC)则能反映机体总抗氧化的能力。GAA对猪抗氧化性能和糖代谢关键酶的影响见表4。

表4 GAA对猪抗氧化性能和糖代谢关键酶的影响
GAA通过增强抗氧化酶活力,增强机体抗氧化能力。而GAA的抗氧化能力可能与肌酸水平有关。Lawler等[22]通过体外试验发现,高浓度肌酸能够清除体内自由基。有报道称,磷酸肌酸可以降低MDA含量,提高SOD和CAT活性,从而减轻阿霉素造成的氧化应激,起到保护心肌的目的[42]。Maddock等[43]研究发现,当发生氧化应激损伤后,可通过肌酸增强DNA活性,保证其正常功能。
血液中酶活性的变化可以反映机体的生理状态[44]。在生成葡萄糖-6-磷酸和果糖-1,6-二磷酸的过程中,己糖激酶(Hexokinase, HK)催化葡萄糖磷酸化和磷酸果糖激酶(PFK)催化果糖-6-磷酸化分别会消耗1个ATP。三羧酸循环发生在线粒体中,其中异柠檬酸氧化脱羧转化成α-酮戊二酸,这是异柠檬酸脱氢酶(IDH)催化三羧酸循环产生的第1个脱羧反应,此过程伴有NADH的形成[45]。
三羧酸循环生成的NADH能够直接进入呼吸链,与氧气结合发生反应,产生2.5个ATP。胍基乙酸可以增加NADH-辅酶Q还原酶和ATP合成酶的活性,这也许和肌酸水平增加有关。磷酸肌酸能催化线粒体中ATP的形成,保持能量正常运转[46-47]。ATP水平的增加可以促进能量储存组织如肌肉和脂肪的合成。肌酸是胍基乙酸通过肝脏和SAM形成的,磷酸肌酸和ADP则是肌酸经肌酸激酶(CK)催化再与ATP结合形成的,ATP消耗量低于ATP合成量,因此在饲粮中添加GAA,可增加ATP水平。
4 小结
饲粮中添加胍基乙酸,不仅能够增加畜禽饲料利用率,调节能量代谢,改善肉品质,而且还可以增加养殖经济效益。作为新型的饲料添加剂,胍基乙酸在我国养殖业的应用当前还处在摸索时期,对胍基乙酸的最佳添加剂量、耐受性和作用机制需要进一步研究和探索。
