西藏麦地卡湿地不同水文期原生动物优势种生态位及其种间联结性*
2021-04-02安瑞志乔楠茜汤秋月
安瑞志 张 鹏 达 珍 乔楠茜 汤秋月 巴 桑
(西藏大学理学院青藏高原湿地与流域生态实验室 拉萨 850000)
原生动物是单细胞真核生物一大类群的统称(Pawlowski, 2014),包括纤毛虫、肉足虫、异养鞭毛虫及具有内共生藻类或功能性叶绿体而进行兼性营养的植鞭毛虫等,是水生态系统微生物食物网、物质循环和能量流动中的重要结构和功能组成部分(沈韫芬,1990;许木启等, 2001)。因其体积小、生命周期短,对全球环境变化和分布反应迅速,在水环境监测方面拥有独特的优势(Xuetal.,2012;杨欣兰等, 2019)。优势种是群落中占优势的物种,是研究生态过程的关键(Avolioetal., 2019)。生态位是反映物种在群落和生态系统中的位置和状况的重要指标,其研究有助于理解不同物种之间的竞争与共存关系(武承旭等, 2020;安瑞志等, 2020)。种间关系是水生生物群落的本质特征之一,尤其竞争与捕食是驱动群落结构变化的重要因素,对于理解水生生物群落演替、结构和功能具有重要作用(赵萍萍等, 2012;胡梦红等, 2014)。迄今有关浮游植物(吴佳梦等, 2019;杨文焕等, 2020)、游泳动物(Ofomataetal., 2010;邓小艳等, 2018)的研究较多。然而很少有人将上述优势种生态位及其种间联结性分析方法应用于对原生动物群落的相关研究。
青藏高原湿地分布广泛,湿地面积达13.19万km2,占全国湿地面积的20%(邢宇等, 2009;赵志龙等, 2014)。青藏高原的大部分湿地受人类活动影响较小,保持着相对稳定的原始生态系统,因而备受国内外研究生物起源、进化、区系和迁徙的学者广泛关注(安瑞志等, 2021;蔡倩倩等, 2013; Zhangetal., 2018; Guetal., 2019)。麦地卡湿地是拉萨河的河源地区(中国科学院青藏高原综合科学考察队,1981),是西藏3个被列入国际重要湿地名录的湿地之一。该湿地在调节区域气候、水资源供应和保持生物多样性等方面具有重要的生态安全屏障作用。目前,有关麦地卡的生物学研究文章仅限于种子植物(拦继酒等, 2018),有关原生动物的研究鲜见报道。
为了探究西藏麦地卡湿地国家级自然保护区不同水文期原生动物群落优势种生态位及种间联结性,笔者于2018年7月(丰水期)和10月(枯水期)在麦地卡湿地原生动物进行了定性和定量采集,共采集水样168个,主要探讨原生动物群落优势种是否具有明显的时空变化,从生态位和种间联结性角度对该湿地的群落演替、种间关系等开展研究,揭示不同水文期原生动物群落优势种的资源利用状况、功能地位及生态适应性等相关信息,以期为进一步研究麦地卡湿地原生动物群落的稳定性和空间分布格局提供依据。
1 研究区概况
研究区位于麦地卡湿地国家级自然保护区(30°51′04″—31°09′44″N,92°45′55″—93°19′25″E),地处西藏自治区那曲市嘉黎县麦地卡乡境内,湿地总面积88 052.37 hm2,属高原亚寒带半湿润气候区,区域内只有冷暖季节之分,冷季(冬春)寒冷而风大,年平均气温0.9 ℃,年日照时数为2 211.8 h;年均降水量694 mm,多集中于6—9月;年蒸发量1 400 mm;年无霜期78天(马广仁, 2016)。地貌为山原宽谷盆地,海拔4 570~5 736 m,海拔高差986 m,平均海拔4 900 m(中国科学院青藏高原综合科学考察队,1983)。湿地类型齐全,功能完善,是高寒湖泊、河流与沼泽湿地生态系统中的典型代表,是研究高原湖泊湿地生态、河流湿地生态以及沼泽湿地生态系统的理想场所(王恒颖, 2014)。
2 研究方法
2.1 样点设置
根据麦地卡湿地的生态环境特征、功能分区与梯度结构等特征,设置了14个采样点(图1),分别于2018年7月(丰水期)和10月(枯水期)进行原生动物样品和水体理化因子调查。各样点的基本信息见表1,其中pH、盐度(Salt)、水温(water temperature, WT)和溶解氧(dissolved oxygen,DO)为2次采样期间的平均值。
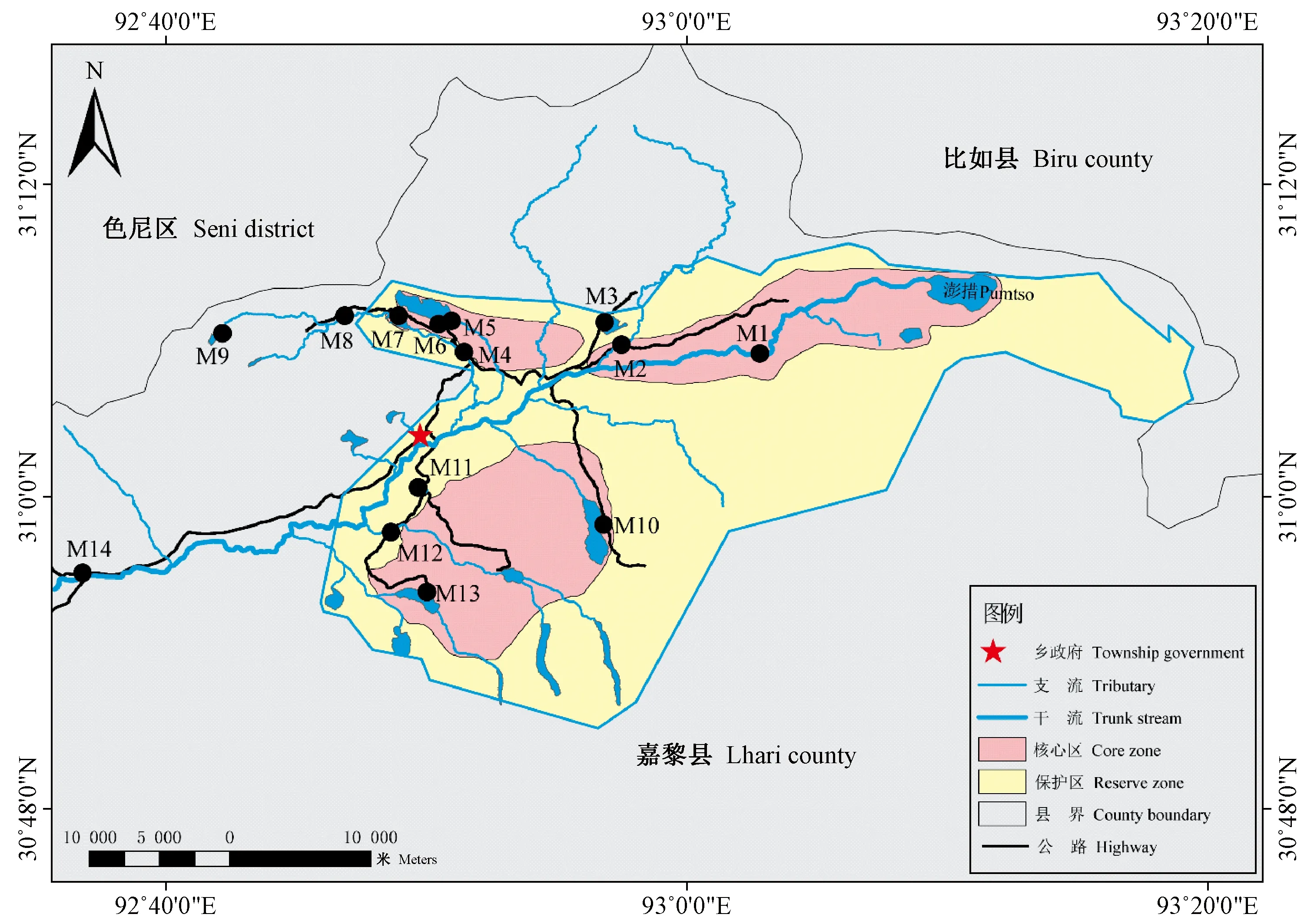
图1 麦地卡湿地样点分布Fig. 1 Sampling sites in the Mitika Wetland
2.2 样品采集和分析
原生动物根据《淡水浮游生物研究方法》(章宗涉等,1991)中规定的方法进行采集和鉴定。定性样品用25#(孔径64 μm)浮游生物网在0~0.5 m处作“∞”形捞取8~10 min,将浓缩样品装入标本瓶, 在室内利用Olympus CX23研究型光学显微镜,采用活体镜检和固定染色法鉴定物种(中国科学院青藏高原综合科学考察队,1983;沈韫芬,1999;周凤霞等, 2011)。定量样品在0~0.5 m处采集1 L混合水样,随后立即用鲁哥氏碘液固定,样品带回实验室沉淀48 h后浓缩至30 mL,移取0.1 mL浓缩匀液至0.1 mL计数框,在10×40倍显微镜下全片计数,每个样品计数2~3次。
现场使用PCSTestr35防水型多参数笔试测试仪(EUTECH,美国)测定水体pH、水温、电导率(electrical conductivity,EC)、盐度及总溶解盐(total dissolved salts,TDS), 使用HI98193微电脑溶解氧测定仪(HANNA,意大利)测得溶解氧, 使用HI98703微电脑多量程浊度测定仪(HANNA,意大利)测得浊度(turbidity,TUR)等,并分别进行3个平行检测。严格按照GB3838-2002《地表水环境的质量标准》对水样进行采集、保存与运输,送有检测资质公司对总磷(total phosphorus,TP)、总氮(total nitrogen,TN)、氨氮(NH3-N)、硝酸盐氮(NO3-N) 和重铬酸盐指数(dichromate oxidizability,CODCr)等指标进行测定。
2.3 数据处理
2.3.1 优势种 依据McNaughton优势度指数(Y)确定优势种(Mcnaughton,1967; Habibetal.,1997),计算公式如下:
(1)
式中,fi为第i种出现的频率,n为同一样点中个体总数,ni为第i种的个体数;Y>0.02则为优势种,Y>0.1为绝对优势种。
2.3.2 生态位 生态位宽度(Bi)采用Colwell等(1971)加权修正的Levins指数,计算公式如下:
(2)
式中,Bi为物种的生态位宽度,Pij为物种i在第j个样点的个体数占该种所有个体数的比例,Pij=Nij/Ni,i表示物种,j为资源,Nij为种群i利用资源状态j的数量,Ni为种群i的总数量,r为采样点数。
生态位重叠(Oik)指数选用Pinaka指数(Piankaetal.,1973),计算公式如下:
(3)
(4)
(5)

2.3.3 种间联结性 为了对麦地卡湿地2个水文期原生动物群落优势种的种间联结性进行分析, 本研究采用方差比率法(variance ratio ,VR)对总体关联性进行测定,结合χ2检验定性判定种间联结性,并以联结系数(association coefficient ,AC)和共同出现百分率(percentage co-occurrence,PC)为辅助参数, 并经过点相关系数(φ)的检验测定, 以增加结果的精确性和可信度(程瑞梅等, 2013;董静瑞等, 2017;戴冬旭等, 2019)。总体关联性用方差比率法(Schluter,1984)计算,计算公式如下:
(6)
(7)
(8)
(9)
W=N·VR。
(10)

种间联结性检验采用χ2值通过连续校正公式(张金屯, 2004)计算如下:
(11)
式中,a为2个物种组成的种对同时存在的样点之和,b和c为其中仅出现一个物种的样点之和,d为一对物种共同出现频率为0的样点数,因χ2计算结果无法反映负值,笔者利用ad-bc的符号对物种间的正、负联结性进行判断,若其值为正数,可判定为正联结,反之则判定为负联结,相等则判定为无联结;若χ2<3.841(P>0.05),表示物种间无显著联结性,若3.841≤χ2≤6.635(0.01≤P≤0.05),表示物种间联结显著,若χ2>6.635(P<0.01),则表示物种间联结性极显著。
在χ2检验的基础上, 结合联结系数(AC)和共同出现百分率(PC)来分析物种不仅能反映种间联结的真实情况, 而且可以进一步验证χ2检验中不显著的种对间的联结性,从而对这部分种对的联结性进行具体测度,得到更加精确的结果。AC可以测定种间联结程度(张金屯, 2004),计算公式如下:
(12)
(13)
(14)
式中,联结系数AC取值区间为[-1,1],如果AC=0,说明物种间完全独立,AC值越趋向1,表示物种间正联结程度越高,AC值越趋向-1,则物种间的负联结性越强。
采用PC分析物种间正联结程度(张金屯, 2004),计算公式如下:
(15)
式中,PC取值区间为[0,1],当PC取值越偏向1,表示物种间联结程度越强,当物种间联结性不存在时,PC值等于0。
在种间联结性中,AC和PC能体现出那些由χ2检验判断为不显著种对的联结性及大小,但PC夸大了2个物种均出现时种间联结显著程度,AC夸大2个物种均不出现时的联结显著程度, 而点相关系数(φ)可以降低这2种情况的影响(潘琪等, 2015)。φ用于解释种间联结的程度(张金屯, 2004),计算公式如下:
(16)
式中,φ取值在[-1,1]范围内有效;φ值越偏向1,物种间的正联结性越强;φ值越偏向-1,物种间的负联结性越强。
利用ArcGIS对调查区域经纬度数据进行可视化标注处理;通过Excel 2016软件计算优势度指数、生态位宽度指数、生态位重叠指数和种间联结性。
3 结果与分析
3.1 优势种
通过定性和定量样本综合分析,丰水期共鉴定出原生动物141种,枯水期为138种,其中优势种9种,隶属2门5属。倒卵形隐滴虫(Cryptomonasobovata)和球形砂壳虫(Difflugiaglobulosa)是2个水情期共有优势种,优势种出现频率、丰度和优势度在不同水文期均有差异(表2)。丰水期优势种较少,仅有4种,波豆虫属(Bodosp.)的丰度和优势度最高,为丰水期绝对优势种(Y>0.1);枯水期优势种较多,为7种,卵形隐滴虫(C.ovata)和球形砂壳虫为绝对优势种(Y>0.1)。
3.2 生态位
3.2.1 生态位宽度 麦地卡湿地丰水期和枯水期原生动物优势种的生态位宽度如表3所示。其中,丰水期优势种的生态位宽度变化范围为[0.101,0.445],生态位宽度最大的是波豆虫属(0.445),其次是球形砂壳虫(0.376),生态位宽度最小的是小茄壳虫(Hyalospheniaminuta,0.101);枯水期优势种的生态位宽度变化范围为[0.141,0.272],生态位宽度最大的是球形砂壳虫(0.272),其次是吻状隐滴虫(C.rostrate, 0.269),生态位宽度最小的是钟形钟虫(Vorticellacampanula, 0.141)。根据各优势种在不同水情期的生态位宽度将优势种分为3类。第一类的生态位宽度值较大(Bi≥0.25),为广生态位的种类,包含丰水期的波豆虫属、枯水期的吻状隐滴虫及2个水情期均为优势种的球形砂壳虫等3种,此类优势种对环境的适应性能力强,具有较广的生态适应幅度;第二类的生态位宽度值较小(0.15≤Bi<0.25),为中生态位的种类,包含枯水期的卵形隐滴虫、蛋白核隐滴虫(C.pyrenoidifera)、回转隐滴虫(C.reflexa)和倒卵形隐滴虫等4种,这一类对生境有一定要求,对环境资源具有较强的选择性,在适宜的条件下能够充分利用资源;第三类的生态位宽度值小(0≤Bi<0.15),为窄生态位的种类,包含丰水期的倒卵形隐滴虫、小茄壳虫和枯水期的钟形钟虫等3种,这一类对环境因子具有一定的依赖性,对水环境条件变化比较敏感,适应性较弱。

表2 麦地卡湿地不同水文期原生动物优势种出现频率(fi)、丰度(A) 及优势度(Y)①Tab.2 Frequency (fi), abundance (A) and dominance (Y) of dominant protozoan species under different hydrologic periods in the Mitika Wetland

表3 麦地卡湿地不同水文期原生动物优势种生态位宽度(Bi)Tab.3 Niche breadth (Bi) of dominant protozoan species under different hydrologic periods in the Mitika Wetland
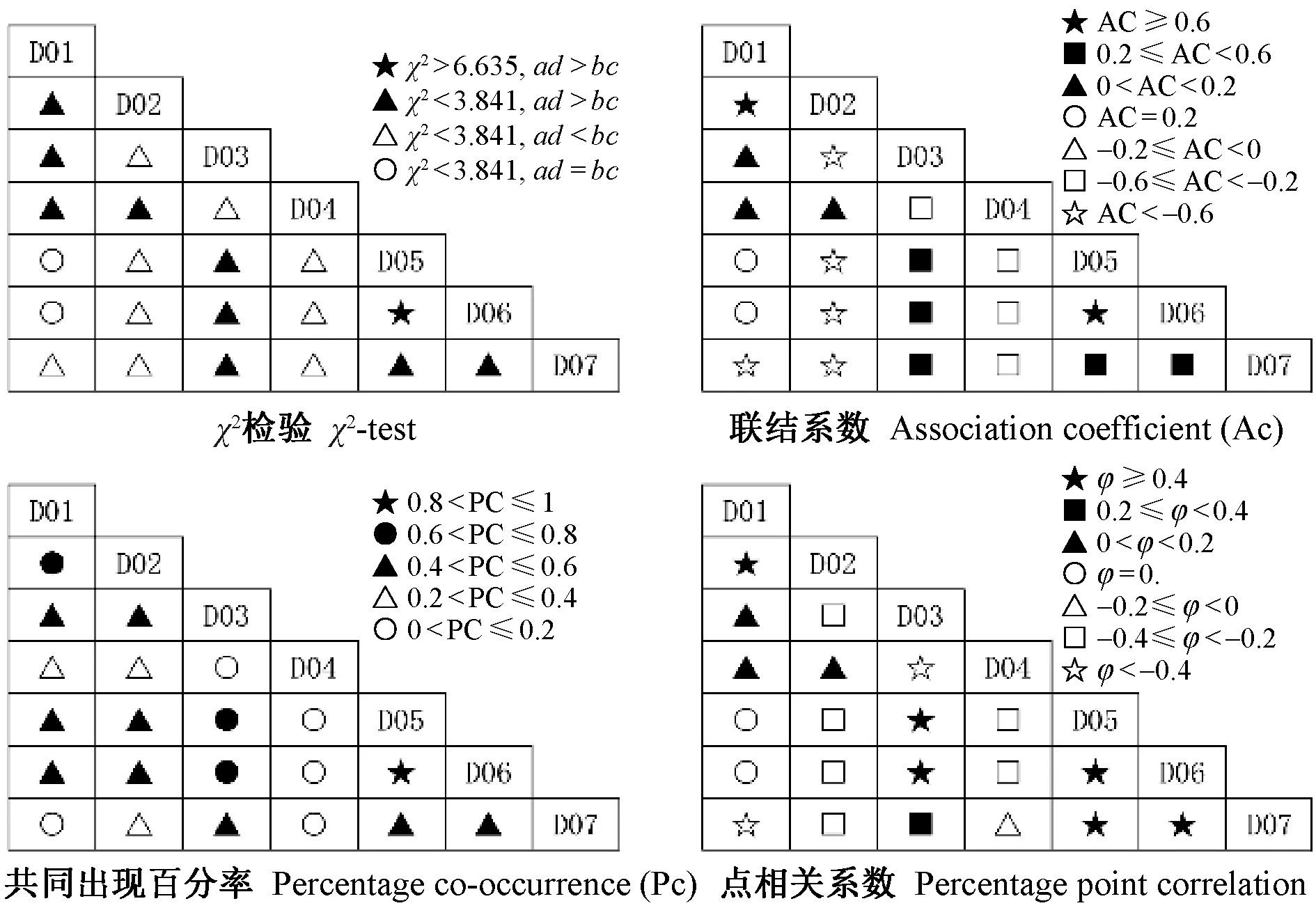
3.2.2 生态位重叠值 麦地卡湿地不同水文期原生动物优势种生态位重叠值如表4和表5所示。2个水情期物种间对资源利用或生态适应能力的相似程度存在较大差异性,说明不同优势种之间,对环境资源的利用既相互独立,又相互重叠。2个水文期9个优势种的生态位重叠值变化范围为[0.001,1]。其中,丰水期的生态位重叠值变化范围为[0.089,0.456](表4),未出现重叠度较高(Oik>0.6)的种对,2对重叠度较低(0.3 3.2.3 生态响应速率 麦地卡湿地不同水情期原生动物群落优势种的相对资源占有量和生态响应速率结果如表6所示。可以看出,丰水期以小茄壳虫的相对资源占有量的变化幅度(即绝对值)最大,枯水期以钟形钟虫的相对资源占有量的变化幅度最大。丰水期,小茄壳虫有最大的负ΔOik,表明该种衰退空间最大,倒卵形隐滴虫同属于衰退型,生长空间同呈缩小趋势;波豆虫属和球形砂壳虫处于发展型,其中球形砂壳虫有最大的正ΔOik,表明该种发展空间最大,与其他优势种在资源利用上竞争激烈,且占据竞争优势;枯水期,球形砂壳虫、卵形隐滴虫和钟形钟虫属于衰退型,其他各优势种处于发展型,蛋白核隐滴虫和回转隐滴虫有最大的发展空间,均大于其他优势种;吻状隐滴虫的发展空间略小于蛋白核隐滴虫和回转隐滴虫,倒卵形隐滴虫发展空间最小。 基于生态响应速率可进一步确定不同水文期优势种对生境条件的生态响应状况。丰水期,波豆虫属的发展趋势较为强烈(R=1.072),球形砂壳虫的发展趋势较弱(R=0.277);倒卵形隐滴虫的衰退趋势较为强烈(R=-0.423),小茄壳虫的衰退趋势较弱(R=-0.069)。枯水期,倒卵形隐滴虫的发展趋势较为强烈(R=0.062),蛋白核隐滴虫和回转隐滴虫的发展趋势较弱(R=0.030);球形砂壳虫的衰退趋势较为强烈(R=-0.049),钟形钟虫的衰退趋势较弱(R=-0.013)。从总体上看,2个水文期生态响应速率之和均为正数,表明麦地卡湿地原生动物群落处于发展阶段,尚未演替至顶级阶段。 表4 麦地卡湿地丰水期原生动物优势种生态位重叠值(Oik)①Tab.4 Niche overlap value (Oik) of dominant protozoan species in the Mitika Wetland during the wet season 表5 麦地卡湿地枯水期原生动物优势种生态位重叠值(Oik)Tab.5 Niche overlap value (Oik) of dominant protozoan species in the Mitika Wetland during the dry season 表6 麦地卡湿地不同水文期原生动物优势种相对占用资源量(ΔOik)及生态响应速率(R)Tab.6 Relative occupation resource quantity(ΔOik)and ecological response rate(R) of dominant protozoan species under different hydrologic periods in the Mitika Wetland 表7 麦地卡湿地不同水文期原生动物优势种的种间总体关联性Tab.7 General inter-specific association of dominant protozoan species under different hydrologic periods in the Mitika Wetland 3.3.2 种间联结测度 麦地卡湿地不同水文期原生动物群落优势种的χ2检验、联结系数、共同出现百分率和点相关系数结果如图2、3所示。 丰水期,χ2检验结果显示,在6个种对内,均为联结性不显著χ2<3.841(P>0.05)种对,未出现显著或者极显著联结种对;正联结性3对,负联结性3对,群落正负联结比为1,表明群落结构相对稳定,正联结性的物种互利共生,负联结性的物种种间竞争和排斥作用较强,物种间对资源环境的需要和适应不尽相同,处于相对稳定的演替中期阶段。联结系数结果显示,正联结对数为3对,其中有1对正联结系数程度较高(AC≥0.6),约占总对数的16.67%; 1对正联结系数程度一般(0.2≤AC<0.6),约占总对数的16.67%;1对正联结系数程度趋向独立(0 枯水期,χ2检验结果显示,在21个种对内,联结性不显著χ2<3.841(P>0.05)的种对有20对,占21个种对的95.24%;联结性极显著χ2>6.635(P<0.01)的种对有1对,占21个种对的4.76%;正联结性10对,负联结性9对,无联结性2对,群落正负联结比为1.11,表明群落结构稳定,物种间稳定共存,群落发育成熟,处于稳定的演替末期阶段。其中,正联结极显著χ2>6.635(P<0.01,ad>bc)1对,为蛋白核隐滴虫和回转隐滴虫形成的种对。联结系数结果显示,正联结对数为10对,其中有2对正联结系数程度较高(AC≥0.6),约占总对数的9.52%;5对正联结系数程度一般(0.2≤AC<0.6),约占总对数的23.81%;3对正联结系数程度趋向独立(0 图2 麦地卡湿地丰水期原生动物优势种的种间联结性Fig. 2 Interspecific association of dominant protozoan species in the Mitika Wetland during the wet season 图3 麦地卡湿地枯水期原生动物优势种的种间联结性Fig. 3 Interspecific association of dominant protozoan species in the Mitika Wetland during the dry season 生态位宽度反映了物种对资源利用、分布状况和均匀程度,是衡量物种对其生境利用程度(李德志等, 2006;李显森, 2013)。麦地卡湿地原生动物优势种由广生态位种、中生态位种和窄生态位种组成, 并以广生态位种占主导地位, 这与张琳琳等(2020)和樊紫薇等(2020)的研究基本一致。其中,球形砂壳虫为广生态位种,在麦地卡湿地2个水情期均为优势种,表明球形砂壳虫数量较多,出现频率较高,这主要是因为球形砂壳虫属于有壳肉足虫,最显著的特征是变形体外面被包一外壳,对虫体具有保护作用(张文静, 2010),表现出较强的生态适应性;而窄生态位种则分布空间狭小,持续时间较短,如小茄壳虫仅在丰水期成为优势种,对水环境条件变化比较敏感,表现出较弱的生态适应性。因此,生态位宽度较高的优势种在麦地卡湿地分布较广,且数量较多;而生态位较窄的优势种仅在某一季节成为优势种或在部分样点数量较多,这与吴佳梦等(2019)和杨文焕等(2020)的研究基本一致。 生态位重叠值反映了物种间对资源利用或环境适应能力的相似性特征和竞争关系(王祥福等, 2008)。麦地卡湿地2个水文期原生动物群落优势种重叠值低(0 种群呈发展或衰退趋势是不同种群竞争的直接结果,是生物与非生物因子的相互作用的生态响应(孟东平等, 2006)。本研究中,从原生动物优势种相对资源占有量(ΔOik)来看,较大生态位宽度的原生动物优势种,往往具有较广的资源利用谱,且大多数呈较强的发展空间, 如丰水期的波豆虫属、枯水期的吻状隐滴虫以及2个水文期均为优势种的球形砂壳虫等;而具有较小生态位宽度的原生动物优势种,对水环境的适应性较差,大多数属于衰退型种群,如丰水期的小茄壳虫和枯水期的钟形钟虫等。从原生动物优势种的生态响应速率来看,发展趋势较强优势种为了获得更多的资源,可能会通过增大生态位宽度来加强自身的生态适应型,将会逐渐代替其他衰退型的原生动物,如丰水期的球形砂壳虫和枯水期的吻状隐滴虫。而衰退趋势较强的原生动物,如丰水期的小茄壳虫和枯水期的钟形钟虫等将会逐渐消亡。同时也应该看到,枯水期生态位宽度最大的球形砂壳虫呈现最为严重的衰退趋势,可能的原因是球形砂壳虫分布范围达到了峰值,随着环境条件的变化,由于自疏或他疏作用使其种群数量增长缓慢甚至有所减少,表现出明显的衰退趋势,这与冯宜明等(2018)和孙延军等(2019)的研究结果较一致。 种间联结性大小反映群落中种间的相互作用和群落演替,总体联结性反映了群落的稳定性(刘润红, 2020)。本研究中,麦地卡湿地2个水文期的原生动物群落优势种总体关联性呈显著正关联,表明该湿地原生动物物种组成和优势种等群落结构较为稳定且存在正向演替的趋势,这与吴佳梦等(2019)和刘润红等(2020)的研究结果较一致。χ2检验判断了麦地卡湿地原生动物群落丰水期4个优势种和枯水期7个优势种的种间联结显著程度。从整体上看,麦地卡湿地原生动物群落的正负联结比排序为:枯水期(1.11)>丰水期(1),表明麦地卡湿地原生动物群落从丰水期至枯水期经历了中期(相对稳定)至末期(稳定)的阶段。但大多数种对的关联性未达到显著水平,表明丰水期和枯水期优势种种对间联结性不强,种间关系较为松散,可能与物种的生物学特性、空间分布的异质性及多样性有关,这与戴冬旭等(2019)和刘惠等(2019)的研究结果较一致。原生动物个体较小,营浮游生活,生命周期短,繁殖快,对环境变化敏感,种群大小时空变化快(沈韫芬,1999),在生境中相互依赖程度低,在时间空间上各自占据有利自身的位置,从而导致原生动物群落中物种优势度对物种关联的影响也并不显著。同时χ2检验并未发现极显著负联结种对,说明他们种间竞争力并不激烈,这可能是由于这些优势种生境存在一定差异,并且群落物种组成又较为复杂,2个物种相遇的概率就越低,在一定程度上导致部分种对的联结性偏弱,因此出现极显著负联结为0的情况(戴冬旭等, 2019)。 联结系数(AC)结果显示,丰水期,1对正联结系数程度较高(AC≥0.6),1对负联结系数程度较高(AC<-0.6);枯水期,2对正联结系数程度较高(AC≥0.6),5对负联结系数程度较高(AC<-0.6)。这个结果是对χ2检验中不显著的种间联结性的进一步验证,表明大部分优势种具有相似的生物学特性或生境需求,并且生态位重叠值也较高,这与李超男等(2017)和董静瑞等(2017)的研究结果基本一致。根据共同出现百分率(PC)可知, 联结程度最强的种对仅有1对,为枯水期蛋白核隐滴虫和回转隐滴虫形成的种对,均属隐滴虫属,二者样点分布、出现频率、生态位宽度等一致,能够资源共享,生态位重叠值较高,可能存在竞争关系(何友均等, 2006)。经过点相关系数的测定,如枯水期的球形砂壳虫与卵形隐滴虫的联结系数与共同出现百分率均有明显降低, 因此经过点相关系数测定后的种间联结性可以更加准确地判定麦地卡湿地原生动物的种间关系,表明点相关系数可以对χ2检验、联结系数和共同出现百分率测定结果进行修正,使物种联结性研究结果更加精确(潘琪等, 2015)。同时点相关系数也表明麦地卡湿地2个水文期的物种间总体趋于正相关,这与χ2检验、联结系数和共同出现百分率结论相一致。 在麦地卡湿地原生动物群落优势种中,丰水期的波豆虫属、枯水期的吻状隐滴虫以及2个水文期均为优势种的球形砂壳虫等对环境的适应性能力强,具有较广的生态适应幅度;而丰水期的小茄壳虫和枯水期的钟形钟虫等对水环境条件变化比较敏感,适应性较弱;部分优势种种间生态位重叠与宽度特征并不一致,如枯水期的蛋白核隐滴虫和回转隐滴虫的重叠值高但生态位宽度较小; 麦地卡湿地原生动物群落物种组成和优势种等群落结构较为稳定且存在正向演替的趋势,但优势种之间以及各优势种与环境之间未达到相对的动态平衡,原生动物群落仍处于发展阶段,尚未演替至顶级阶段,还需从时间、空间和时空等维度进一步研究不同时空维度原生动物资源分配的驱动机制问题。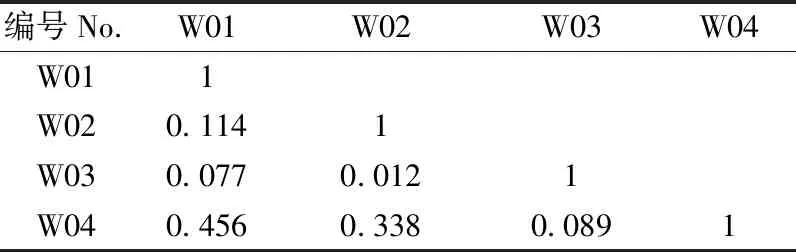
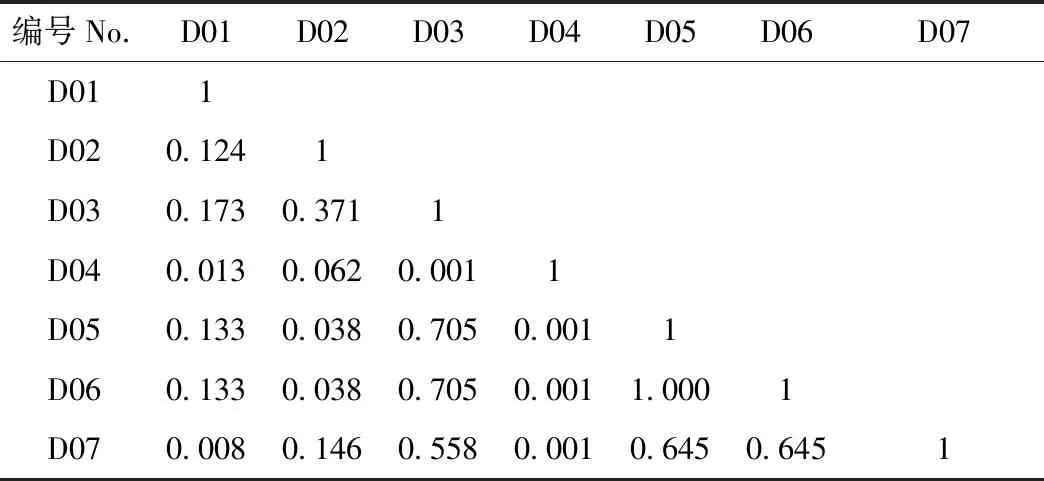
3.3 种间联结性




4 讨论
4.1 优势种生态位
4.2 种间联结性
5 结论
