小叶黄杨越冬叶片呈色及其温光影响因子*
2021-04-02彭金根姜雪茹谢利娟
彭金根 姜雪茹 谢利娟 刘 燕
(1.深圳职业技术学院应用化学与生物技术学院 深圳 518055; 2.北京林业大学园林学院 北京 100083;3.江西农业大学林学院/园林与艺术学院 南昌 330045)
常绿植物冬季叶片发生颜色改变是一种普遍发生的现象(Hughes,2011)。北京地区常见常绿园林树种统计表明,29种常绿树中有21种为针叶树,而阔叶树只有8种(赵芒,2012),然而,常绿阔叶树种越冬进程中叶片还时常发生变红现象。调查发现,生长于荫蔽环境下的黄杨(Buxussp.)、扶芳藤(Euonymusfortunei)、蚊母树(Distyliumracemosum)等叶片能保持绿色或者接近于绿色,而生长于曝光环境下的同种植株叶片则出现程度不一的褐化现象(马海慧,2004)。黄杨属中的小叶黄杨‘丽城’(Buxusmicrophylla‘Belvedere’)植株在冬季不同冠层会呈现亮红色、红棕色、棕色、绿棕色或绿色(姜雪茹等,2015),该现象也表明常绿阔叶植物冬季叶片不同呈色与叶片所处的温光环境差异密切相关。
关于影响常绿阔叶树种冬季叶片呈色因子,有研究认为常绿树种冬季叶片变红表明植物遭受了光氧化伤害(董丽, 2001; 马海慧, 2004),而Peng等(2016)对小叶黄杨(Buxusmicrophylla)研究认为叶片冬季变红是植物的一种适应性表现,关于常绿树种冬季叶片中花色素苷合成功能的研究支持了该观点(Zhuetal.,2018, Zhangetal.,2019)。锦熟黄杨(Buxussempervirens)叶片在冬季光照条件下由绿色转变为红色,与其越冬期间叶片中叶绿素a、b的大量降解以及脱水花菱草类黄素(anhydro-eschscholtzxanthin,一种红色类胡萝卜素)的合成有关(Idaetal., 1995; Hormaetxeetal., 2004)。同一植株不同冠层位置,冬季锦熟黄杨叶片会呈现绿色、棕褐色、黄色、橙色和红色的差异,这与叶片的受光角度不同有关(Hormaetxeetal., 2005)。此外,Hughes等(2005)对常绿阔叶植物加莱克斯草(Galaxurceolata)的报道表明越冬期间叶片变红与其合成花色素苷有关,且变红程度与光照强度正相关,Yu等(2020)对夏季6种天然林优势树种的研究进一步证实叶片中花色素苷合成量与环境中光照强度的高低呈正比关系。
常绿植物冬季叶片变红与冬季低温和高光环境密不可分在不少研究中均得到证实,但针对叶片变红的温度、光照因子的量化研究却未见报道。本研究以北京地区露地栽植的3个小叶黄杨(B.microphylla)品种为研究对象,连续2年对越冬返青期间叶片的呈色情况和栽植地的温度和光照进行追踪测定,对影响小叶黄杨越冬叶片变红的温、光因子进行量化,以期为北方冬季常绿阔叶树种在园林中的应用和提升绿色景观提供理论依据。
1 研究区概况
本研究在北京植物园(116°19′E、40°01′N)内进行,小叶黄杨种植于公园内苗圃。北京的气候类型为北温带半湿润大陆性季风气候,四季分明,春秋两季比较短,夏季高温降雨量多,冬季寒冷较为干燥。试验期间,植株的平均株高×冠幅为1.3 m×1.2 m×1.2 m,所有植株均呈东西向排列,平行种植于空旷的苗圃地。
2 材料与方法
2.1 试验材料
试验材料为北京植物园科研中心从国外引进的3个小叶黄杨品种原株,其中‘绿美’(‘Greenbeauty’)为1997引自美国,‘阳光’(‘Sunnyside’)为1998年引自荷兰,而‘冬绿’(‘Wintergreen’)为2002年引自比利时,在植物园露天苗圃地已栽植近20年,植株生长良好。
2.2 试验方法
2.2.1 植株遮光处理 对3个小叶黄杨品种植株进行连续2个越冬返青期(2011—2013年)进行遮光处理。Lobos等(2012)的研究表明,黑色遮荫网不会改变光质(光谱成分),只改变照光强度。在小叶黄杨植株上方约20 cm处用黑色遮荫网覆盖,覆盖时叶片为正常的绿色(分别为2011及2012年10月中旬)。
2011—2012年,设置4个遮光梯度,分别为:1)全光下阳生叶,100% SUL; 2)透光度为50%~60%的阳生叶,50%~60% SSUL; 3)透光度为30%~40%的阳生叶,30%~40% SSUL; 4)透光度为1%~10%的阴生叶,1%~10% SHL。2012—2013年在前1年的基础上增加“透光度为10%~20%的阳生叶”,即10%~20% SSUL的处理。
小叶黄杨植株5个不同处理下的光照值采用Licor-250光量子计进行测定,测定时间为搭遮荫网后晴朗无云日的中午,测定位置为遮荫网下方15 cm左右,光合有效辐射(photosynthetic active radiation, PAR)值为5个处理下6~8个点的平均值。不同处理水平的划分依据为处理下光照与全光下光照的相对辐射量变化,其中:相对辐射量=1-遮光下PAR/全光下PAR×100%。
2.2.2 温度及光照条件测定 1)越冬返青期间温度测定 采用温度数据记录器U23-004(Onset HOBO公司),对小叶黄杨种植地2011—2013年(当年10月至次年3月)气温实时监测数据每小时自动采集1次。
2)越冬返青期间总太阳辐射值测定 2011—2013连续2个越冬返青期间种植地的日累计太阳辐射总量值取来自小叶黄杨种植地附近的北京气象局气象站(39°48′N,116°28′E; ID号:54511)。
2.2.3 叶片呈红指数(RI)测定 在室内散射光下,用相机(Panasonic Lumix DMC-LX5)拍摄5种不同光照强度处理下叶片,每个处理采集5片叶子,叶片采集方法见彭金根等(2011)。叶片颜色定量分析采用开源软件Fiji (Schindelinetal.,2012, http:∥fiji.sc/)。叶片的照片导入软件后,软件将叶片呈色情况以红(R)、绿(G)、蓝(B)3种颜色的直方图呈现,并标识出各种颜色所占的百分数值。小叶黄杨叶片的变红程度用呈红指数(Redness Index,简称RI)来表示,RI=R/(R+G+B)。
2.2.4 叶片红色色素分布观测 对3个小叶黄杨品种新鲜的全光阳生叶(100%SUL)、遮光阳生叶(50%~60%SSUL,30%~40%SSUL,10%~20%SSUL)及阴生叶(1%~10%SHL)作徒手切片,观测叶片中红色色素的分布状况,并进行拍照,采样时间分别为2012年10月20日和2013年1月22日。显微镜摄像机(LEICA DM500-ICC50 HD,Leica Microsystems, Wetzlar, Germany)成像,目镜的放大倍数为4,照片的像素分辨率为2 048 ×1 536。通过Photoshop CS3(Adobe Systems, San Jose, CA, USA)对切片照片进行亮度和图像清晰度的调整。
2.3 数据处理与分析
采用Excel进行数据处理,采用SPSS 18. 0软件(SPSS Inc., Chicago, USA)进行分析。利用单因素方差分析进行方差检验,多重比较采用Tukey HSD-test(其中显著性P≤ 0.05)。
3 结果与分析
3.1 越冬返青期间小叶黄杨种植地日均温及日太阳总辐射变化
由图1A、1B可知,小叶黄杨种植地试验期间的日均温和太阳总辐射总体表现为随着越冬的深入而呈下降趋势,随着返青的来临而呈上升趋势,其中日太阳总辐射表现出更大波动。 2011—2012年最冷月的日平均温较2012—2013年更高。为探明气温与太阳辐射对叶片呈色的影响,本研究进一步分析了试验期间采样前5日的日均温及太阳辐射值,由图1C、1D可知,采样前5日日均温和平均太阳辐射在不同越冬年份间表现出差异,其中2011—2012年,1月出现日均温最低值,12月出现平均太阳辐射最低值,而在2012—2013年试验期间,12月出现采样前5日日均温的最低值,而12月和1月在平均太阳辐射最低值上差异不显著。此外,2011—2012年和2012—2013年2个越冬返青期,太阳总辐射量在返青后的3月仍处于上升趋势。
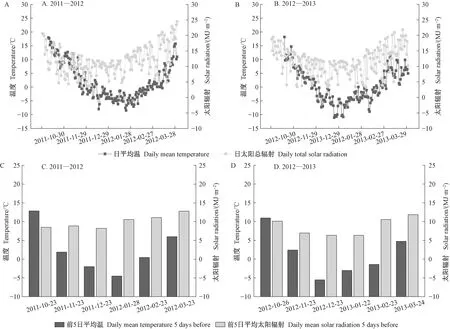
图1 越冬进程中种植地日均温、日太阳总辐射(A:2011—2012,B:2012—2013)及采样前5日平均温、平均太阳辐射(C:2011—2012,D:2012—2013)Fig. 1 The daily temperature and total solar radiation of the planting fields during the overwintering period and the average daily temperature and solar radiation 5 days before sampling
3.2 越冬返青过程中3个小叶黄杨品种叶片呈色变化
3.2.1 不同遮光度下3个品种阳生叶和阴生叶的呈色观测情况 3个小叶黄杨品种不同遮光度下阳生叶在越冬返青期间表现出较大的呈色差异(图2),其中小叶黄杨‘绿美’的变红程度最低,当其叶片处于50%~60%全光时基本能保持绿色; 小叶黄杨‘阳光’的变红程度属于中等水平,当光照强度下降至全光的30%~40%时能保持绿色; 小叶黄杨‘冬绿’的变红程度最为严重,只有阴生叶(光强为全光的1%~10%)才呈现绿色。同时期相比,小叶黄杨叶片远轴侧(即背面)较叶片近轴侧(即正面)变红程度更深。

图2 3个小叶黄杨品种不同光照条件下越冬返青期间叶片呈色情况(2011—2012年)Fig. 2 Leaf color of three cultivars of Buxus microphylla under different light conditions during the period of overwintering and regreening (2011-2012) SUL:阳生叶Sun leaves; SSUL:遮光阳生叶Shading sun leaves; SHL:阴生叶Shade leaves。下同。The same below.
2012—2013年越冬返青期间,试验增设了10%~20%SSUL的遮光处理(图3),结果进一步证实,当环境中光照强度下降至全光的50%~60%时,小叶黄杨‘绿美’就能保持绿色,而小叶黄杨‘阳光’需要环境光下降至全光的30%~40%,小叶黄杨‘冬绿’需要下降至全光的10%~20%。同样,3个小叶黄杨品种同时期叶片远轴侧比近轴侧变红程度更深。无论是2011—2012年还是2012—2013年越冬返青期,光照强度仅为全光1%~10%的3个小叶黄杨品种阴生叶(SHL)整个过程均能保持绿色。

图3 3个小叶黄杨品种不同光照条件下越冬返青期间叶片呈色情况(2012—2013)Fig. 3 Leaf color of three cultivars of B. microphylla under different light conditions during the period of overwintering and regreening (2012—2013)
3.2.2 3个小叶黄杨品种叶片呈红指数(RI)值监测 本研究对越冬返青期间3个小叶黄杨品种叶片的呈红指标(RI)进行了追踪监测(表1),并通过该值与叶片所处环境中的温度和太阳辐射值的对比分析,2011—2012年越冬返青过程中,3个品种的RI值均随着越冬进程中气温的下降呈上升趋势,而随着返青时气温的回升而有所下降。3个品种均在2012年1月28日叶片变红程度最严重,且叶片的远轴侧值较近轴侧值更高。从遮光处理的效果看,所有品种叶片的RI值均表现为随着环境中光照强度的降低而下降。
2012—2013年越冬期间,3个小叶黄杨品种叶片的RI值监测结果见表2。与2011—2012年相比,2012—2013年增设了10%~20%的遮光处理,2个越冬返青期的结果表现相似,3个品种叶片的RI值均随越冬返青期间气温的下降而上升,随气温的上升而下降。此外,2012—2013年期间3个品种叶片在气温最低的1月份RI值也表现最高,小叶黄杨‘绿美’、‘阳光’和‘冬绿’分别是0.384、0.400和0.406。
3.3 越冬返青期间3个小叶黄杨品种叶片呈色与温光因子相关性分析
3.3.1 3个小叶黄杨品种叶片呈红指数RI与温光因子相关性分析 为揭示越冬返青期间气温变化对小叶黄杨品种叶片呈色的影响,本试验分析了越冬返青期间3个品种叶片呈红指标RI值与叶片采样前5日均温的线性回归系数,由表3可知,3个品种叶片越冬返青期间RI值无论是叶片近轴侧还是远轴侧,与气温的回归系数R2均达到显著水平,其中全光下的阳生叶(100% SUL)和遮光50%~60%处理下的阳生叶(50%~60% SSUL)的相关性均达到极显著水平(P≤ 0.001)。
为进一步分析越冬返青期间光照强度变化对小叶黄杨品种叶片呈色的影响,本试验也对小叶黄杨品种叶片RI值与采样前5日平均太阳辐射值进行了线性回归相关系数分析,结果如表4所示,2个越冬返青期(2011—2012年和2012—2013年)该相关性表现出差异,其中在2011—2012年越冬期间,不同遮光处理下的3个品种叶片与采样前5日平均太阳辐射表现出很弱的相关,但2012—2013年越冬期间,两者则表现出较强的相关,具体表现为该越冬返青期,大部分处理的叶片和大多数时期的RI值与采样前5日平均太阳辐射具有显著相关性,2012—2013年越冬期间品种种植地的太阳辐射变化与气温变化具有更高的协同性(图1)可能造成不同年份差异。
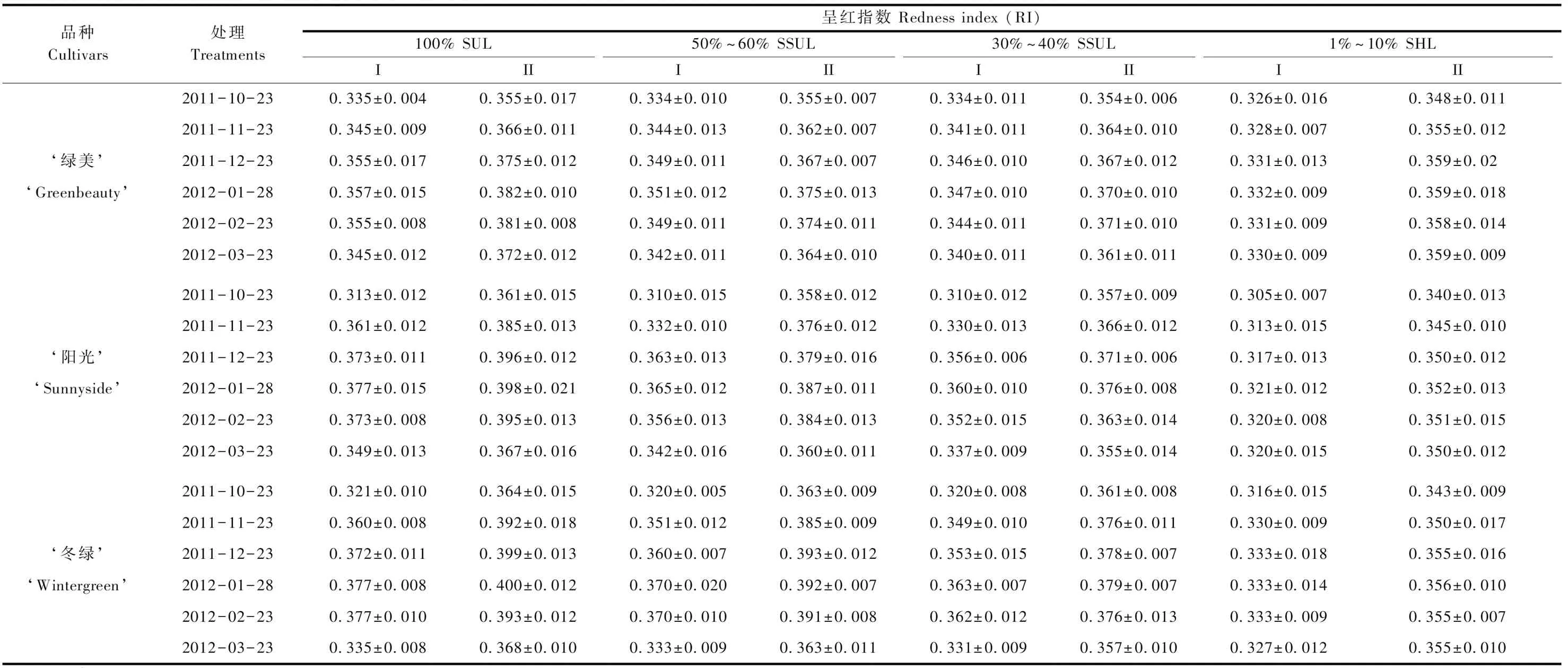
表1 越冬返青期间3个小叶黄杨品种叶片呈红指数(RI)值①Tab.1 The leaf redness index (RI) of three cultivars of B. microphylla during the overwintering and regreening period 2011—2012

表2 越冬返青期间3个小叶黄杨品种叶片呈红指数(RI)值Tab.2 The leaf redness index (RI) of three cultivars of B. microphylla during the overwintering and regreening period 2012—2013

表3 3个小叶黄杨品种叶片呈红指数RI与采样前5日均温线性回归相关系数(R2)①Tab.3 Correlation coefficient (R2) between leaf redness index (RI) and mean temperature of 5 days before sampling of three cultivars of B. microphylla
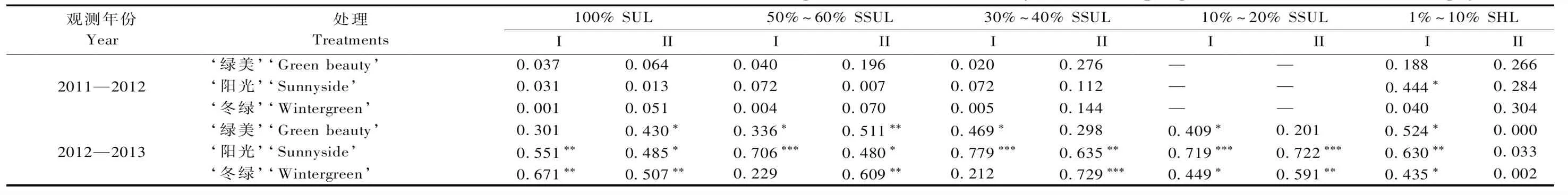
表4 3个小叶黄杨品种叶片呈红指数RI与采样前5日平均太阳辐射线性回归相关系数(R2)Tab.4 Correlation coefficient (R2) between leaf redness index (RI) and the average solar radiation of 5 days before sampling of three cultivars of B. microphylla
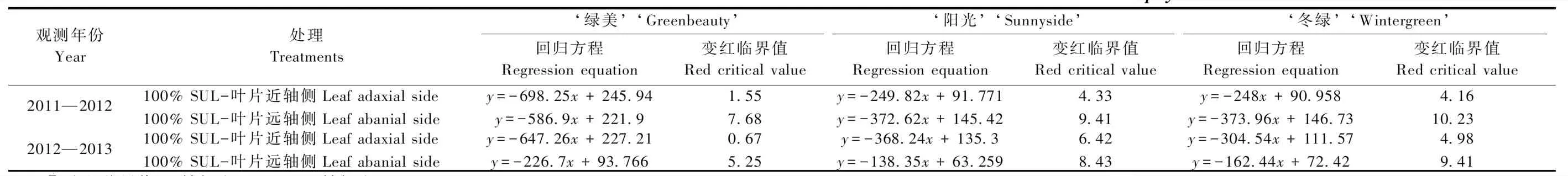
表5 影响小叶黄杨越冬叶片变红的临界日均温分析①Tab.5 Analysis of critical daily mean temperature affecting the leaf reddening of B. microphylla

表6 影响小叶黄杨越冬叶片变红的临界前5日平均太阳辐射分析Tab.6 Analysis of critical solar radiation 5 days before sampling affecting the leaf reddening of B. microphylla
3.3.2 小叶黄杨越冬叶片变红的临界温光因子 根据3个小叶黄杨品种叶片近轴侧和远轴侧2011—2013连续2个越冬时期的呈色表现(图2,图3)和同时期相对应的叶片呈红指标RI值(表1,表2),将叶片近轴侧变红的临界值取值为RI =0.35,而叶片远轴侧变红的临界值取值为RI=0.365。为研究影响3个小叶黄杨品种叶片变红的临界温度值,进行了呈红指标与温度值的回归方程拟合分析,3个品种的变红临界温度值均具有差异性,且不同年份间也表现出差异(表5)。2011—2012年和2012—2013年越冬返青期间,变红程度轻保绿效果好的‘绿美’变红临界温度值最低,其中近轴侧为1.55 ℃和0.67 ℃,远轴侧为7.68 ℃和5.25 ℃; 变红程度处于中间水平且返青较晚的‘阳光’,近轴侧为4.33 ℃和6.42 ℃,远轴侧为9.41 ℃和8.43 ℃; 变红程度最高但返青早的‘冬绿’,近轴侧为4.16 ℃和4.98 ℃,远轴侧为10.23 ℃和9.41 ℃。
另外,本试验也对影响小叶黄杨品种冬季叶片变红的采样前5日的平均太阳辐射临界值进行了分析,3个小叶黄杨品种越冬叶片在该值上未表现出品种差异(表6),2011—2012年和2012—2013年2个越冬时期,影响3个小叶黄杨品种冬季叶片变红的临界太阳辐射值基本都在10 MJ·m-2左右,只是2个越冬时期间稍有差异。
3.3.3 小叶黄杨品种越冬前后叶片红色色素分布变化 本试验通过观测不同光照条件下3个小叶黄杨品种越冬前后的叶片的徒手切片显微观察(图4A~E),研究3个品种叶片中红色色素的分布差异。3个品种无论是全光阳生叶(SUL)还是遮光阳生叶(SSUL),越冬过程中红色色素积累的部位均为叶片上层的栅栏组织,且红色色素分布范围随着光照强度的增加而增加。不同品种表现一定差异,变红程度最轻的小叶黄杨‘绿美’,当环境中光照强度为全光的50%~60%时,红色色素的积累量非常少(图4B);变红程度中等的‘阳光’,当光照强度降低至全光的30%~40%时红色色素积累量几乎看不到(图4D);而对于变红程度最严重的‘冬绿’来说,即便当光照强度降低全光的10%~20%,最冷月(1月)时,仍有少量红色色素积累(图4F)。
4 讨论
4.1 影响叶片呈色的光温因子
部分常绿植物在冬季叶片发生颜色改变是一种普遍发生的现象(Hughes,2011),常绿植物越冬期间的这种变红通常只发生在气温偏低的冬季且存在照光的条件下,处于温暖季节或者处于遮荫条件下则很少发生变红(Hormaetxeetal., 2004; 彭金根等, 2011),也有报道指出在夏季极端高温胁迫下锦熟黄杨的叶片发生变红(Hormaetxeetal., 2007)的特殊现象。本文对3个具有不同越冬呈色表现的小叶黄杨品种的叶片呈色进行连续2个越冬返青时期(2011—2013年)的跟踪测定,同时对该时期植株栽植地的气温和太阳辐射值进行连续跟踪记录,最终得到影响3个品种叶片越冬变红的临界气温值和临界太阳辐射值。
冬季的低温不影响常绿植物叶片对光能的捕获,但会导致植株叶片光合作用中参与卡尔文循环的酶活性降低(Tanaka, 2007),从而造成光能吸收和光能利用间的不平衡(Öquistetal., 2003; Verhoeven, 2014)。这种持续过剩的光能最终会导致叶片中活性氧的大量合成(Ruellandetal., 2009; 姜雪茹等, 2015)。活性氧能对叶绿素起到漂白作用(Choudhuryetal., 2001),而且在拟南芥(Arabidopsisthaliana)叶片的花色素苷合成中发挥重要的分子调控作用(Vanderauwera, 2005)。3个小叶黄杨品种栽植地的日均温2011—2013年越冬返青期间呈明显的先下降后上升的趋势,而越冬期间(3月前)日太阳辐射值变化幅度小(采样前5日的太阳辐射平均值在10 MJ·m-2上下),说明在越冬进程中叶片发生变红现象,气温的下降可能发挥着更关键作用。
前期的研究发现,小叶黄杨‘丽城’(B.microphylla‘Licheng’)冬季不同光照下植株叶片呈现绿色、绿棕色、棕色、红棕色和亮红色5种不同颜色(姜雪茹等, 2015),而在锦熟黄杨植株上冬季也发现了类似的情况(Hormaetxeetal., 2005)。本文通过对越冬返青期间3个小叶黄杨品种叶片呈红指数RI与环境中温光因子进行线性拟合分析,得出影响小叶黄杨品种叶片近轴侧和远轴侧冬季变红的温光临界值,其中变红程度最低冬季基本保持绿色的‘绿美’、返青最晚而变红程度中等的‘阳光’以及返青最早但变红程度最深的‘冬绿’在2011—2013年越冬时期,叶片近轴侧和远轴侧变红的临界温度具有明显差异,而3个品种越冬期间在影响叶片变红的采样前5日平均太阳辐射总值上均在10 MJ·m-2左右,无明显差异。



图4 3个小叶黄杨品种不同光照条件下越冬前(2012-10-20)、后(2013-01-22)显微结构Fig. 4 Observation hand-sliced leaves of B. microphylla before (2012-10-20) and after(2013-01-22)winter under different light conditionsad:叶片正面Adaxial; ab:叶片背面Abaxial.
综上可知,光温因子对叶片呈色具有直接影响,3个品种叶片冬季变红与越冬返青期间低温和相对强光照的交互作用密切相关,叶片越冬期间在呈色上的品种差异,反映不同品种对低温+高光的敏感程度和耐受性存在差异。除此之外,本试验的研究结果还表明,3个小叶黄杨品种叶片的远轴侧和近轴侧相比,越冬时前者对光照的反应更为快捷,说明两者在越冬返青期间面对低温和高光的双重胁迫可能采取了不同的光响应策略。
4.2 光温因子对小叶黄杨越冬叶片红色色素合成与分布的影响
3个品种在越冬前和越冬后5个光照强度下叶片的切片结构(图4)可知,所有品种变红叶片中红色色素的积累与光照强度呈正相关,且所有品种阳生型叶片红色色素主要分布于接触了直射光的近轴侧上端,这与前人对同属的锦熟黄杨的研究情况相似(Hormaetxeetal., 2005; Silva-Cancinoetal., 2012)。另外,从品种特性上说,3个小叶黄杨品种对照光的敏感程度依次为‘冬绿’、‘阳光’和‘绿美’,这与叶片呈色情况较为一致(图2、图3),同时也与这3个品种冬季阳生叶叶绿素荧光动力学活性相统一(彭金根等,2011)。3个品种变红的叶片中红色色素的分布情况与在小叶黄杨‘丽城’不同树冠部位(光照强度不同)叶片的叶片解剖观察结果相一致(姜雪茹等,2015),但本研究设置了不同遮光梯度,揭示了不同光照强度值对红色色素分布范围的影响。
花叶木藜芦(Leucothoefontanesiana)(Hughes, 2011)、忍冬(Lonicerajaponica)(Carpenteretal., 2014)等植物种类中变红叶片中红色色素分布与黄杨属植物相似。部分研究报道了红色色素同时分布于叶片近轴侧和远轴侧,如加莱克斯草(Hughesetal.,2007)、黧蒴锥(Castanopsisfissa)(Zhangetal. 2016)、青芋(Colocasiaesculenta)(Hughesetal., 2014)、肖蒲桃(Acmenaacuminatissima)(Zhuetal., 2018)等。导致常绿植物叶片冬季变红的红色色素主要是花色素苷及红色类胡萝卜素(彭金根等, 2017),而前者主要存在于液泡中,是具有水溶性的黄酮类物质 (Gould, 2004),而后者主要分布于有色体的悬浮滴状物中,是具有脂溶性的萜类物质(Ytterbergetal., 2006),因而常绿植物叶片中红色色素在叶片中的分布情况,除了受不同程度的照光强度以及不同角度的照光方向的影响外,还与叶片中红色色素的类型密切相关。
5 结论
本文探究影响3个小叶黄杨品种叶片变红的温、光环境量化条件。变红程度最低的‘绿美’其变红临界温度最低,越冬变红程度最深但返青早的‘冬绿’叶片变红临界温度最高;而变红程度中等但返青晚的是‘阳光’该值也处于中间水平。3个品种叶片呈红指数RI均随着光照强度的下降而降低,表明越冬期间小叶黄杨叶片变红与冬季的低温和相对高光的相互作用密切相关。3个品种的阳生叶越冬期间在呈色上表现不同,反映了它们在低温和相对高光双重胁迫环境下的耐受性及敏感程度存在差异。在北方园林应用时,小叶黄杨适宜种植在遮荫环境下,有利于其冬季维持较好的绿色景观。
