毛乌素沙地油蒿光系统II多时间尺度的环境响应特征
2021-04-02李鑫豪徐铭泽高圣杰魏宁宁查天山
靳 川,蒋 燕,李鑫豪,徐铭泽,高圣杰,魏宁宁,贾 昕,田 赟,查天山
(1. 北京林业大学水土保持学院宁夏盐池毛乌素沙地生态系统国家定位观测研究站,北京 100083;2. 北京林业大学水土保持国家林业局重点实验室,北京 100083)
0 引言
作为维持生态系统物质代谢和能量循环的基础,植物光合作用非常容易受到环境波动的影响,不同强度和持续时间的环境胁迫会协同抑制光合过程[1-2]。光系统II(Photosystem II,PSII)能量分配被认为是光合过程中最为敏感的部分,PSII天线色素分子吸收光能有3个去向:1)进行光化学反应,通过碳同化合成有机物[3];2)通过叶黄素循环以热能形式将过剩光能耗散[4-5];3)重新释放一个光子,即产生叶绿素荧光[6-7]。3种途径相互竞争权衡,且受光合有效辐射(Photosynthetically Active Radiation,PAR)、空气温度(Ta)、饱和水汽压差(Vapor Pressure Difference,VPD)和土壤含水率(Soil Water Content,SWC)等环境因素影响[5,8-9]。上述环境变量通过各自波动周期、频率和振幅调控植物PSII能量分配[10-11]。在分-秒时间尺度上,PAR增加激活叶黄素循环,植物通过改变调节性热耗散(ΦNPQ)和非调节性热耗散(ΦNO)的比例耗散过剩光能[12];在小时时间尺度上,VPD升高可导致蒸腾加速,植物调节气孔减少水分损失[13],同时也不可避免地抑制了实际光化学效率(ΦPSII);在日尺度上,由PAR、Ta和VPD的天循环驱动PSII能量分配[14];在几天至数月时间周期上,高辐射、寒潮、热浪和降水等天气事件以及植物的物候期都会导致PSII能量分配变化[10,15]。尽管有上述认识,但目前关于PSII能量分配的相关研究大多集中在室内短期控制试验,难以模拟自然中真实的环境动态[16-17]。具有高太阳辐射、极端温度和水文梯度跨度大等特点的荒漠生态系统则更加复杂[18]。荒漠夏季光照远超植物所需,导致光合机构产生氧化损伤[19];极端温度抑制了相关酶的活性,造成光合效率降低[20-21];空气干湿循环引起植物气孔调节[13,22],导致荧光淬灭[23]。另外,荒漠植物也常因长期缺水造成PSII失活,产生可逆的光损伤[20-21,24]。因此,荒漠植物PSII能量分配与各环境因子在多时间尺度上很可能具有明显关联特征。
频谱处理可检验数据序列的时间动态格局[25],如小波分析。小波分析方法已经广泛应用于多个生态相关领域,适用于探究多组时间序列数据之间关系[25-27]。然而,目前几乎没有研究将小波分析应用于叶绿素荧光的长期监测数据上,尤其缺乏植物对环境波动在多时间尺度下响应过程和调节机制的理解认识。因此,本研究对宁夏盐池毛乌素沙地优势荒漠物种油蒿(Artemisia ordosica)进行叶绿素荧光长期原位连续监测,采用小波分析方法探究油蒿PSII能量分配在多时间尺度下的环境响应特征,旨在为荒漠植物对不同环境胁迫的敏感程度、适应能力和生理可塑性提供数据支撑,同时为荒漠地区植被恢复物种选择提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏吴忠市盐池荒漠生态系统定位研究站(37°53"08""N,107°25"46"" E,平均海拔1530 m),是半干旱区和干旱区之间的交错带,由于过度放牧等人为因素该区植被已严重退化,近20年的封育使该区植被得到了有效的恢复,这也被认为是该区荒漠化逆转的重要标志。该区属温带大陆性季风气候,平均气温为8.3 ℃,昼夜温差大,全年昼夜平均温差可达25 ℃以上。年平均潜在蒸散量为2024 mm,年平均降水量为287 mm,降雨稀少且不均匀,主要集中在6—9月,4—5月容易发生春季干旱。土壤以风沙土和黑垆土为主,密度为1.54 g/cm3。盛行西北风,年平均风速2.6 m/s。研究地植被群落以旱生植物为主,优势灌木物种有油蒿(Artemisia ordosica)、沙柳(Salix psammophila)和杨柴(Hedysarum mongolicum),观测期内相对覆盖度分别为53 %、17 %和12 %。
1.2 试验材料
油蒿广泛分布于中国北部及西北部荒漠地区,因其根系发达耐沙埋、耐旱,在固定流沙和促进群落演替中发挥着重要作用[17]。油蒿通常在4月萌发,8月积累生物量达到最大,9月末开始凋落。样地内油蒿平均密度为1.43株/m2,平均高度为34.7 cm,平均冠幅为0.64 m。选取3株长势相同且具有代表性(高度为50 cm,冠幅为0.8 m,约50分枝左右)的油蒿作为样本植株。避免监测期内人为因素的干扰,保证样本植株处于完全自然生长的状态,并进行物候的同步观测。
1.3 叶绿素荧光监测与计算
于2016年4月28日至10月3日,采用多通道连续监测荧光仪(MONITORING-PAM2000,WALZ公司,德国)对油蒿样本植株进行24 h原位连续监测,测量间隔为30 min。用荧光探头的特制叶夹固定叶片,调整探头方位使叶片充分受光,每隔7 d定期检查叶片并及时更换。测定叶绿素荧光参数包括饱和脉冲下荧光(Fm")和实时荧光(Fs)。将夜间的Fm"、Fs分别作为暗处理最大荧光(Fm)和暗处理最小荧光(Fo)。根据Kramer公式计算PSII能量分配[28],实际光化学效率(ΦPSII,%)、调节性热耗散(ΦNPQ,%)和非调节性热耗散(ΦNO,%)的计算方法如下:
式中Fm"为饱和脉冲下荧光,Fs为实时荧光,Fm为暗处理最大荧光。
采用式(4)计算最大光化学效率(Fv/Fm)
式中Fo为暗处理最小荧光。
1.4 环境气象因子监测与计算
光合有效辐射由距荧光仪100 m处通量塔(高度为6 m)配有的辐射传感器(PAR-LITE,Kipp & Zonen公司,荷兰)测得。空气温度和相对湿度由塔上温湿度传感器(HMP155A,Vantaa公司,芬兰)测得,采用式(5)和式(6)[16]计算饱和水汽压差(VPD,kPa)
式中e为饱和空气压力,kPa;RH为相对湿度,%;Ta为空气温度,℃。
土壤含水率由样地内土壤温湿度传感器(ECH2O-5TE,Decagon Devices公司,美国)测得,深度为30 cm。降雨量由距样地200 m处空地内雨量筒(TE525W,Campbell公司,美国)测得。所有环境气象因子测量间隔均为30 min。
1.5 数据处理与分析
本研究使用连续小波变换(Constant Wavelet Transform,CWT)和小波相干性(Wavelet Coherence,WTC)分析荧光参数的时间序列。CWT是将时间序列数据在固定间隔的变量进行平移或伸缩等变换,用母小波来表示。CWT可以反映时间序列数据在不同时间尺度的波动[29],数量为N的离散变量xn的积分谱 ()xnWs定义为
式中δt为时间间隔,为复共轭尺度,s为小波变换尺度,n为数组1至N,n"为正整数取值。本研究选取Morlet小波作为母小波,采用式(8)计算xn的小波功率谱(Sn)[26,30],得到其贡献最大的时间周期
定义2个时间序列数据之间的交叉小波功率谱(Cn),相位角谱(An)和WTC功率谱(Rn2)[26,30]
式中x为环境因子,y为植物荧光参数,为变量x与y的交叉小波变换,S为时间尺度的平滑处理,和分别为的实部和虚部[31](在时频域图中,An可以用箭头来绘制,若x与y正相关,向右的箭头表示x与y的相位是一致的,向左则相反。而向下的箭头表示y滞后于x90°或270°,向上的箭头表示x滞后于y90°或270°。每360°为1个时间周期)。
2 结果与分析
2.1 环境因子与油蒿PSII能量分配参数动态
PAR求日最大值和平均值,Ta求日最大值、日最小值和日平均值,VPD和SWC求日平均值,降雨量求日总值。如图1所示,观测期内PAR和Ta趋势相近,在5 —7月呈上升趋势,8—9月呈下降趋势。PAR最高达1963μmol/(m2·s)(6月8日),7月和8月出现持续多天低值(PAR<400μmol/(m2·s))。Ta最高值达32.1 ℃(7月29日),最低值为2.9 ℃(5月6日),出现1次大幅降温事件(8月22日)。VPD在5—7月呈周期性波动,8—9月较低,日平均值最高达2.5 kPa(7月25日)。SWC整体呈持续下降趋势,在8月20日降雨量达到52.6 mm的降水后有较大增加。如图2所示,ΦPSII在7月和8月有2处明显的下降,日平均值最低为0.45%(7月16日)。ΦNPQ和ΦNO趋势大致相同,7月和8月有2处明显增高。ΦNPQ日平均值最高达23 %(7月16日),ΦNO日平均值最高达36%(7月3日)。Fv/Fm呈周期性波动,最高0.82(5月31日),最低0.68(5月3日)。观测期内,各叶绿素荧光参数和环境因子具有明显季节波动。
PAR、Ta和VPD都呈现先升高后降低的日变化趋势,PAR峰值在13:00左右,Ta和VPD峰值约在16:00。PAR在5—7月较高,Ta在5月和9月较低,VPD在6—7月较高。ΦPSII呈现与ΦNPQ和ΦNO相反的日变化趋势,峰值出现时间与PAR一致。ΦPSII在6—7月较低,ΦNPQ在5 —7月的峰值较高,ΦNO在7月的峰值较高(图3)。
2.2 环境因子与油蒿PSII能量分配参数在各时间周期内变化
除SWC以外,所有变量全局小波功率谱在1 d时间周期均出现强峰值,并且在4~40 d时间周期内分别出现多次振荡。叶绿素荧光参数(ΦPSII、ΦNPQ和ΦNO)峰值相对环境因子(PAR、Ta、VPD和SWC)较小。SWC既无日尺度谱峰,也无季节尺度谱峰。变量PAR、ΦPSII、ΦNPQ和ΦNO在小于1 d的时间周期内出现多次峰值,但其功率远低于1 d时间周期处(图4)。
小波功率谱可以清楚地显示各变量在观测期内的时频域局部特征。在0.5 d时间周期处,PAR出现显著小波功率区域。在1 d时间周期处,PAR、Ta和VPD均出现显著小波功率区域,区域非连续且强烈震荡。Ta在4~32 d周期内、VPD在8~32 d周期以及SWC在1~32 d周期内均出现小波功率值的升高(图5)。在1 d时间周期处,ΦPSII、ΦNPQ和ΦNO均出现强烈震荡的显著区域。ΦPSII在8~32 d周期内、ΦNPQ在4~32 d周期以及ΦNO在16~32 d周期内也呈现小波功率值的升高(图6)。
2.3 油蒿PSII能量分配参数与环境因子的小波相关性
油蒿PSII能量分配参数与环境因子在多个时间尺度上存在显著小波相关区域,分布不均且波动明显。在日尺度上,ΦPSII与PAR、Ta、VPD和SWC小波相关区域相位角分 别 为(12.32±3.56)°、(47.88±21.20)°、(73.25±13.01)°和(183.69±96.81) °,即ΦPSII滞后于PAR、Ta、VPD和SWC分别为(0.82±0.24)、(3.19±1.41)、(4.88±0.87)和(12.25±6.45) h(图7;表1)。ΦNPQ滞后于PAR、Ta、VPD和SWC分别为(0.62±0.27)、(3.45±1.57)、(4.94±2.64)和(11.87±4.98) h(图 8;表1)。ΦNO滞后于PAR、Ta、VPD和SWC分别为(0.72±0.32)、(3.27±1.43)、(3.83±1.69)和(8.07±6.97) h(图 9;表 1)。油蒿PSII能量分配参数(ΦPSII、ΦNPQ和ΦNO)滞后于PAR、Ta、VPD和SWC分别平均为43 min、3.3、4.6和10.72 h。
在季节尺度上,ΦPSII滞后于PAR、Ta、VPD和SWC分 别 为(13.64±5.54)、(9.87±3.47)、(7.68±1.99)和(23.47±12.46) d(图7;表2)。ΦNPQ滞后于PAR、Ta、VPD和SWC分别为(16.05±7.65)、(3.52±2.73)、(9.90±5.64)和(17.89±14.80) d(图8;表2)。ΦNO滞后于PAR、Ta、VPD和SWC分别为(14.44±6.12)、(8.23±4.47)、(8.73±5.05)和(23.73±10.97) d(图9;表2)。油蒿PSII能量分配参数分别滞后于Ta、VPD、PAR和SWC 7.2、8.8、14.7和21.6 d。
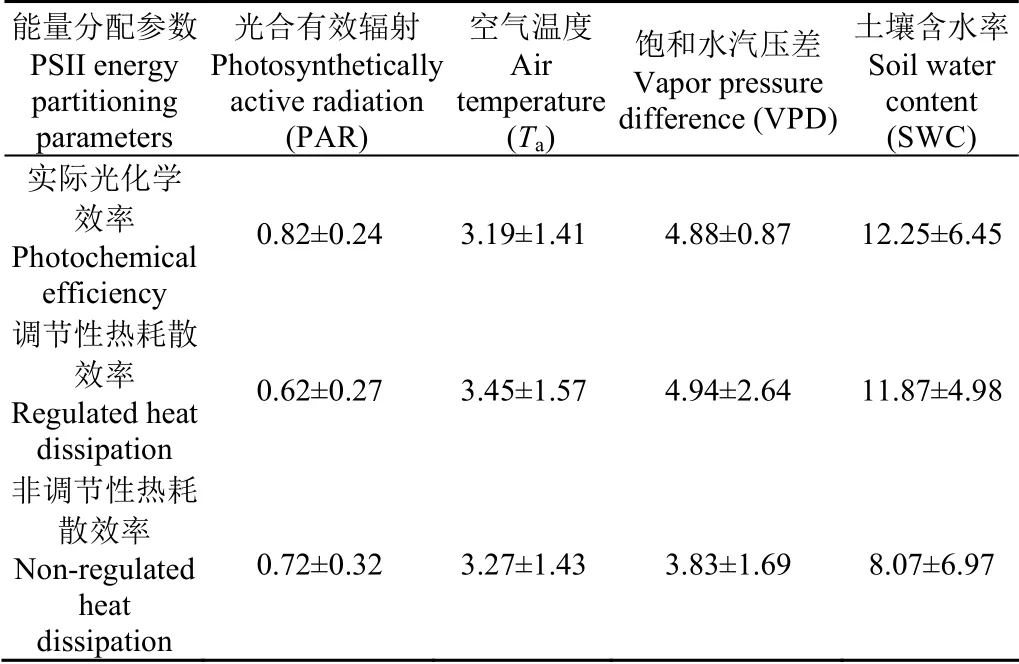
表1 PSII能量分配参数在日尺度上滞后环境因子的时间 Table 1 Lag time of PSII energy partitioning parameters relative to environmental factors on daily scale h
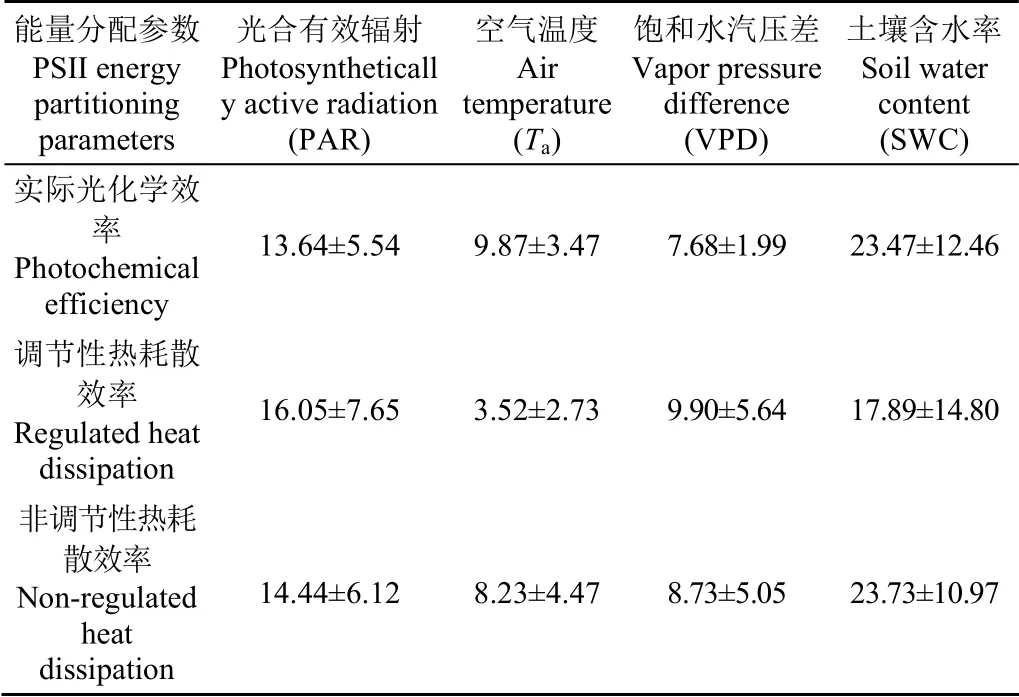
表2 PSII能量分配参数在季节尺度上滞后环境因子的时间 Table 2 Lag time of PSII energy partitioning parameters relative to environmental factors on seasonal scale d
3 讨论
植物通过热耗散机制消耗光合过程中的过剩光能,减少活性氧以避免产生光损伤[23]。将热耗散分为ΦNPQ和ΦNO可获得更精准的定量信息,ΦNPQ表征热耗散等光保护调节机制的强弱[28],ΦNO则指示光抑制或光损伤的程度[10,28]。日尺度上,油蒿PSII能量分配参数与各环境因子均有很强相关性(图7~图9),对PAR变化最敏感且受其主导。PAR增加导致更多过剩光能需要耗散,因此ΦNPQ和ΦNO与PAR正相关,同时不可避免抑制了ΦPSII,这一系列过程是瞬间发生的[5,16]。油蒿ΦPSII在12:00左右发生明显降低,在夜晚恢复(图3)。表明在高辐射下,虽然油蒿光化学反应过程受到了限制,但其PSII反应中心实际并没有产生损害[32-33]。这与同地区不同物种相关研究结果不一致,研究发现沙柳和杨柴因高辐射表现出严重光抑制,并造成永久性光损伤。这可能与植物形态学有关,沙柳叶呈叶线形,杨柴为互生扁平小叶,而具有针形叶的油蒿则有利于避免过量太阳辐射。油蒿PSII能量分配分别滞后Ta和VPD 3.3和4.6 h(表1)。研究表明Ta降低抑制叶绿体捕光蛋白酶活性,从而影响植物光合生理状态[20-21]。清晨ΦPSII下降(图3),此时Ta处于全天最低,油蒿PSII光反应相关酶活性受到抑制,导致ΦPSII早于PAR开始降低[21,34]。VPD增加导致气孔调节,蒸腾速率降低[13,22],抑制了水的裂解[24],也提高了跨类囊体质子梯度[35],最终造成ΦPSII降低,ΦNPQ和ΦNO增加。油蒿PSII能量分配在1 d时间周期内滞后SWC约10.72 h,远高于其他环境因子(表1)。这说明短时间内油蒿对于土壤水的敏感程度较低[10,34]。
荒漠植物需要应对干旱、高辐射和极端温度等多种环境胁迫[5,18,36],PSII长期适应机制的形成需要几天甚至几周时间[10,17],主要通过调整光合色素浓度、叶绿素a/b比例或PSII特定蛋白质含量等途径实现[10,37]。Fv/Fm是表征植物PSII受胁迫程度的关键指标,通常情况下在0.75~0.85之间,发生下降则表示PSII 受损失活[34]。油蒿Fv/Fm和PSII能量分配季节波动明显,但始终维持在一个整体水平上(Fv/Fm约为0.78)(图2)。研究认为油蒿可以通过自身修复调节,使其PSII恢复到正常状态[16,28]。研究发现,油蒿PSII能量分配受物候影响,5月初处于萌芽期,叶绿素含量及活性低,对外界抵抗力较差,易产生光抑制[10,23]。7—8月受高温高辐射影响,产生光抑制造成PSII损伤,此时油蒿处于长势最旺盛的花果期,其生物量和叶片功能性状达到最佳,PSII处于不断受损和修复的过程[32-34]。9月末油蒿叶片老化进入枯萎期,叶绿素含量、各种酶活性伴随光合单元机构衰退降低,低温胁迫导致Fv/Fm低值出现[10,38]。
在季节尺度上,油蒿PSII能量分配滞后Ta和VPD相对PAR和SWC较短。说明在长时间尺度,油蒿PSII对Ta和VPD更为敏感。植物在高VPD下会调节气孔关闭[13,22],增强叶黄素循环等持续性热耗散[23],同时对PSII反应中心造成可逆损害[32-34]。极端温度或高太阳辐射则会打破PSII能量输入和利用的平衡,造成PSII捕光蛋白损伤[39]。与其他研究比较,油蒿对极端温度的适应能力要高于荒漠物种沙柳和红砂(Reaumuria soongorica)[8,36]。荒漠地区昼夜温差大,油蒿可通过提高细胞内渗透压,增加原生质抗凝结性,减缓代谢速率来适应极端温度[39]。油蒿PSII能量分配滞后PAR约14.7 d(表 2),油蒿对于高辐射具有一定的耐受能力[16,34]。干旱是影响植物生长生存的重要环境胁迫,水分限制着细胞内一系列的生理生化反应[13,24]。油蒿具有粗长发达的主根和茂密的侧根[17,40],PSII能量分配滞后SWC时间最长,约21.6 d(表2),可见其对干旱有着极强的耐受性,因此也经常用作干旱区植被恢复的先锋物种。
4 结论
1)日尺度上,油蒿可能通过叶黄素循环等热耗散机制和气孔导度调节PSII能量分配,主要受光合有效辐射(Photosynthetically Active Radiation,PAR)控制,同时受空气温度(Ta)和饱和水汽压差(Vapor Pressure Difference,VPD)的调控,对土壤含水率(Soil Water Content ,SWC)敏感性较差。PSII能量分配参数滞后于PAR 43 min,且分别滞后于Ta、VPD和SWC 3.3、4.6和10.72 h。
2)季节尺度上,油蒿可能通过提高PSII修复能力、增加特定蛋白数量和色素浓度等方式调节PSII能量分配,对Ta和VPD变化较敏感,并且受物候期的影响。PSII能量分配参数分别滞后于Ta、VPD、PAR和SWC 7.2、8.8、14.7和21.6 d。
3)在荒漠恶劣环境下,油蒿可以通过自身调节不断修复受损PSII(Fv/Fm维持在0.78附近),对干旱和高辐射表现出耐受性,具有一定的适应能力和生理可塑性。
