北草蜥(Takydromus septentrionalis)种群间形态与咬合力的差异性分化
2021-03-30梁潇方
梁潇方,计 翔
(南京师范大学生命科学学院,江苏 南京 210023)
温度大小规则(temperature size rule,TSR)是指变温动物在较高温度下发育较快,且在体积较小时性成熟较早的趋势. 体温较低的动物在较高的环境温度下生长得更快,在较小体型时达到性成熟[1]. 然而,和大多数生物学规律都有例外一样,目前,已有大量文献记载的反TSR事例,即在较高的温度下,性成熟个体的体型更大[1-2].
对蜥蜴咬合力功能不同结构组成的研究发现,形态结构之间关系的同时参与了咬合力和咬合持续时间的优化. 即作用于咬合力选择压力的基础是某些形态特征,咬合力增加的代价是咬合力持续时间的减少,反之亦然[3]. 蜥蜴咬合力本身可能受到生存压力和性选择的影响,雄性高咬合力的适应选择可能导致咬合力和头部形状的两性差异[4-6]. 这些过程可能取决于栖息地环境,因此在头部形态两性异形(sexual dimorphis)之间也可能存在趋同进化[7]. 两性异形在爬行动物中很常见[8],在蜥蜴和鳄鱼中,主要是雄性偏多(在长度上雄性比雌性多出50%);而在蛇和海龟中,通常是雌性偏多(在长度上雄性比雌性多出50%).
北草蜥(T.septentrionalis)为中国特有种[9],它们活动在各种大型山脉和水体,分布范围覆盖中国中部和东南部地区[10],是研究种群间性状特征和两性异形进化的理想物种. 目前,关于蜥蜴两性异形的研究主要涉及体型和头部形态,而关于咬合力的研究还未见报道. 本文以北草蜥为研究对象,探讨它们种群间的性状特征(形态和咬合力)是否符合温度大小规则,形态与咬合力之间的关系以及是否存在两性异形现象等.
1 材料与方法
1.1 实验材料
1.1.1 样点的选取
采集动物的5个样点为桂阳(GY:25°75′N,112°73′E)、宁德(ND:26°66′N,119°53′E)、舟山册子岛(ZS:30°05′N,121°56′E)、滁州(CZ:32°29′N,118°31′E)、烟台(YT:37°50′N,121°27′E),依次由低纬度向高纬度逐渐过渡. 同一地区的北草蜥可以看作一个种群,桂阳、宁德、滁州、烟台均为内陆地区,舟山种群所属舟山市位于浙江舟山群岛.
1.1.2 实验动物的采集
选取桂阳(GY)、宁德(ND)、舟山册子岛(ZS)、滁州(CZ)、烟台(YT)5个样点的北草蜥作为实验动物,在2018年和2019年的三月底至四月初从野外徒手抓或用套圈采集北草蜥. 实验动物采集后将雌雄分开饲养,避免在实验开始之前发生交配,以便之后实验的顺利进行. 总计452只(GY:♀93 ♂95,ND:♀21 ♂22,ZS:♀45 ♂47,CZ:♀56 ♂53,YT:♀6 ♂14)北草蜥用于本实验.

A:设计图;B:实物;C:蜥蜴咬合片;D:咬合力峰值图图1 咬合力传感器Fig.1 Bite force sensor design
1.1.3 实验仪器
形态数据使用Mitutoyo数显游标卡尺进行测定,精度为0.01 mm,称量体重使用Mettler电子天平,精度为1 mg.
咬合力仪器为定制咬合力传感器Ⅱ型(量程:100 N,分辨率:0.01 N,重量:500 g,南京神源生智能科技有限公司)测量蜥蜴咬合力(图1 A、B). 在每个咬合片的外表面的末端粘合一条皮革(厚度1 mm)来制备咬合点从而保护蜥蜴的牙齿免受损伤[11].
1.2 实验方法
1.2.1 实验动物的饲养
将采集到的动物带回南京师范大学两栖爬行动物实验室,剪趾标记,以便识别. 置于90 cm×60 cm×50 cm(长×宽×高)饲养缸内饲养,下铺5 cm厚适当湿度的营养土、椰土,放置瓦片和新鲜草皮,种植麦冬供北草蜥攀附. 将具有交配斑的成年雌性和雄性分别置于同样规格的饲养缸内,不同种群蜥蜴分开饲养,避免产生混乱,影响后续实验的进行. 饲养缸置于温度为(28±2)℃的模拟外界环境的玻璃控温室内,缸一端悬挂一只75 W全光谱灯,光照时间段设置为07:00~17:00(阳光充足时关闭全光谱灯,使蜥蜴接受阳光照射,避免眼睛因长时间直射光照受到损伤),灯开启后可在缸内形成室温至55 ℃ 的连续温度梯度,供蜥蜴自主调温. 饲养缸一侧放置培养皿,作为食皿和水皿供蜥蜴自主进食和饮水,每天喂食一次黄粉虫,一周喂食一次白蟋蟀(幼体),定期喂食维生素、钙粉;对感冒的个体中止实验进行隔离(防止交叉感染),同时每日喂食3次含有电解质、维生素和钙粉的水,直至恢复正常;不定期对饲养缸内土壤进行松动,避免因阴暗潮湿滋生细菌,保持饲养环境清洁卫生.
1.2.2 测量形态指标
2019年4月中下旬所有动物采集完毕,在南京实验室对成年个体进行性别识别,称量,记录断尾情况和测量时间,测量内容包括个体的头体长(snout-vent length,SVL)、腹长(abdomen length,AL)、头长(head length,HL)、头宽(head width,HW)、头高(head hight,HH)、张口尺寸(mouth opening,MO[12])、尾长(tail long,TL)、体重(body mass,BM).
1.2.3 测量咬合力
在进行咬力测试之前,将蜥蜴置于一个饲养箱中,在全光谱灯下持续2 h的光照以达到蜥蜴近似的活动体温. 对个体进行3次测试,3次测试之间至少相隔0.5 h,使用RDS软件(用于传感器配置和传感器数据采集)记录每次测试过程中获得的咬合力最大值;在繁殖期间每隔一个月(4月15日,5月15日,6月15日)测量实验个体的形态数据和咬力数据,最后取3次测量均值作为个体本次繁殖季节咬合力大小进行数据分析,避免个体形态和性状随时间发生变化而产生误差.
1.2.4 头部尺寸与咬合力的关系
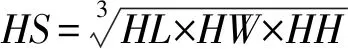
1.2.5 统计分析
采用Bartlett检验和Kolmogorov-Smirnor检验对所有的数据进行方差均质性和正态性检测,使用 STATISTICA 8.0和SPSS 20.0进行统计学分析. 描述性统计值均用平均值±标准误(Mean±SE)来表示,显著性水平均设置为α=0.05. 采用单因素方差分析(one-way ANOVA),多重比较(Tukey’sposthoctest)和主成分分析(principal component analysis,PCA),对形态指标进行描述性统计分析,并对北草蜥不同种群的雌雄间主要形态指标进行比较.
2 结果与讨论
2.1 北草蜥种群间的性状比较
年均温和年降水量(数据来源
种群间形态指标描述统计值(表1)分析显示,SVL(F4,448=94.50,P<0.001)、AL(F4,448=67.78,P<0.001)、HH(F4,448=22.17,P<0.001)和HS(F4,448=63.10,P<0.001)随纬度的升高而逐渐增加,即GY 图2 北草蜥采样点及年均温和年降水量Fig.2 Sampling sites and their annual average temperature and precipitation of T.septentrionalis 表1 北草蜥不同种群身体指标的描述性统计值Table 1 Descriptive statistics of body indicators in different populations of T.septentrionalis 种群间形态多重比较检验(Tukey’sposthoctest)结果显示:(1)SVL在桂阳、宁德和舟山种群与滁州和烟台种群之间差异显著,而滁州与烟台种群间没有差异;(2)HS在桂阳和宁德种群间没有差异,舟山、滁州和烟台种群间也没有差异,但桂阳和宁德种群与舟山、滁州和桂阳种群之间差异显著;(3)BF在桂阳和宁德种群间没有差异,舟山和滁州种群间也没有差异,但桂阳和宁德种群与舟山和滁州种群以及烟台种群之间存在显著差异(表1). 本研究5个样点中,桂阳、宁德、滁州、烟台4个种群所处陆地,且纬度递增,环境温度递减. 研究结果显示北草蜥陆地种群随着所在地区年均温的降低,蜥蜴的形态和咬合力呈现递增的趋势. 但与猜想不同的是,舟山种群的北草蜥表现出不同于其他种群的性状,舟山种群在头长、头宽、张口尺寸、尾长、体重、和咬合力上都大于滁州种群,滁州的年均温小于舟山,而且在地理分布上纬度大于舟山. 这说明在研究结果中环境温度的变化对不同种群的北草蜥形态和咬合力的改变有着重要作用,但不是唯一决定因素,其性状的变化是为了适应当地环境而做出的可塑性突变. 以头体长作为协变量进行方差分析发现,5个种群间咬合力之间没有差异(F4,439=0.867,P=0.484). 以头部尺寸作为协变量分析发现,5个种群间咬合力呈差异显著(F4,439=11.704,P<0.001). 对头部尺寸与咬合力进行回归分析和单因素方差分析(F1,447=1230.26,P<0.001,均质性检验:P<0.001,平行性检验:P<0.001)显示,这两个变量间呈显著正相关(F1,448=1 230.27,P<0.001),对不同种群间的两个变量进行回归分析时,同样具有显著正相关(P<0.001),其回归方程斜率大小为GY A:5个种群所有个体的头部尺寸与咬合力均值之间的回归分析;B:不同种群间北草蜥头部尺寸与咬合力之间的回归分析.图3 北草蜥头部尺寸与咬合力的回归关系Fig.3 Regression relationship between head size and bite force 图4 北草蜥性状指标主成分分析Fig.4 PCA analysis of character indice of the T.septentrionalis 对北草蜥性状特征进行主成分分析选取出主要的指标. 对主成分分析中的第一主成分(PC1)系数较高的指标进行两性之间的比较. 第一主成分的值与各个指标呈正相关,占据74.212%的变异,SVL、HL、HW、HH、MO、HS、BM、BF具有较高的相关系数;第二主成分(PC2)的值与SVL、AL、TL、BM呈正相关,而与HL、HW、HH、MO、HS、BF呈负相关,但相关系数较低,占据10.689%的变异;每个形态指标的分布由95%置信椭圆表示(图4). 两性异形比较分析结果显示,雌雄个体的HS、MO、BF在5个种群中均具有显著差异(allP<0.001),并且雄性的HS、MO和BF均大于雌性;雌性个体的SVL在桂阳、滁州种群中具有差异(P<0.05),但在宁德、舟山、烟台种群中并没有差异,并且除宁德外的4个种群中雌性的SVL都大于雄性;雌雄个体的BM在5个种群中都没有差异,但除了桂阳种群之外的4个种群中雄性的体重都大于雌性(表2). 探究物种性状在种群间的变化是理解物种适应当地生态环境条件必不可少的环节. 已有证据表明,随着个体所处环境温度的升高,体细胞生长和个体发育的速度都在增加,在较高的温度下,达到性成熟的时间更短[13]. 热环境差异是导致个体大小产生地理差异的重要原因之一. 生活在欧洲西南部和非洲西北部潮湿的灌木丛中的阿尔及利亚砂蜥蜴(Psammodromusalgirus)的体型随着海拔的升高而增大,降温速率随着海拔和体型的增大而减小[14]. 岛屿的特殊气候和饮食条件很有可能是造成舟山种群形态和咬合力不同于其他种群的原因. 饮食生态位的差异导致了斯基罗斯岛壁虎(Podarcisgaigeae)的体型、头部形状和进食习惯的差异,有以下4个原因:首先,体型较大的岛屿种群的头部相对较宽. 第二,岛屿蜥蜴进化出更强壮的头部肌肉来应对当地的坚硬猎物[15],因此咬合力也更强[16]. 第三,更大的体型和变异的形态导致了更强的咬合力;这些变异可能是由于基因突变,也可能是适应环境的可塑性反应[17]. 第四,与陆地蜥蜴的猎物相比,岛屿上的硬质猎物比例更高,岛屿蜥蜴吃掉的硬质猎物比例也要大得多[18]. 这就可以解释为何岛屿种群与陆地种群在形态和咬合力之间的差异. 目前国内鲜有关于蜥蜴咬合力的研究,通过对北草蜥的形态与咬合力之间的关系进行探讨,结果表明北草蜥的头部形态与咬合力之间存在显著正相关. 北草蜥雌雄之间在头部尺寸、张口尺寸和咬合力上具有显著差异,即存在明显两性异形现象. 北草蜥两性异形产生的原因可能是饮食(生存压力)和配偶竞争(性选择压力)导致蜥蜴需要进化出更大的张口以咬住猎物,更大的头部来支撑头部肌肉和更强的咬合力以撕裂猎物[19-20]. 雄性间在冲突中使用的武器通常是由性选择驱动进化的,雌性倾向于选择具有“优质”武器的雄性[20]. 同时推测咬合力也可能是雌性对雄性能力的一种预估[21-22]. 综上所述,北草蜥种群间形态和咬合力具有显著差异,而且内陆种群的性状随着所在地区环境温度的降低而增大,岛屿种群则不同,这与岛屿的特殊气候和饮食条件有关;北草蜥的头部尺寸、张口尺寸、咬合力具有明显的两性异形,头部尺寸与咬合力呈正相关. 北草蜥的性状进化形成的差异性与自然环境因素密切相关,这为理解蜥蜴形态、功能和生态之间的关系提供了依据.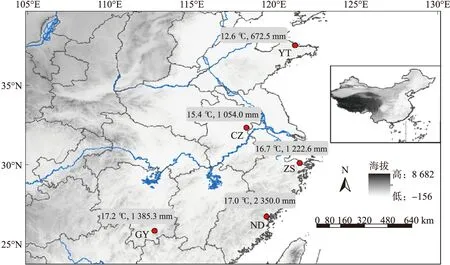

2.2 北草蜥咬合力与头部尺寸的关系


2.3 北草蜥性状指标的两性异形分析
3 结论
