德汉劳绵蟹(Lauridromia dehaani)线粒体基因组测定及短尾下目系统发生分析
2021-03-30邢雨辉邹永梅姚文佳张晨岭印学晨邹修文陈建琴
邢雨辉,周 豪,邹永梅,姚文佳,张晨岭,印学晨,邹修文,陈建琴
(1.南京师范大学中北学院,江苏 镇江 212310)(2.江苏第二师范学院生命科学与化学化工学院,江苏 南京 210013)(3.南京师范大学生命科学学院,江苏 南京 210023)
短尾下目(Brachyura)是现存甲壳动物中物种多样性最高类群之一,广泛分布在世界各个地区的淡水、海洋和陆地中,种类繁多,超过7 250种[1]. 短尾下目的演化历程可看作是辐射演化的实例,在进化的历程中由于不同的生存环境而朝着不同的方向(淡水、海洋和潮间带)发生适应演化,最终形成如今具有极高多样性的类群[2-3]. 正确地认识物种的分类地位和构建稳健的系统发生关系对追溯短尾下目的起源和演化历程具有重要的作用. 早期,Guinot对短尾下目进行分类梳理时,把蟹类两性生殖孔的位置作为高级阶元的分类依据,将短尾下目(Brachyura)分为肢孔派(Podotremata)、异孔派(Heterotremata)和胸孔派(Thoracotremata)[4-5].在Guinot的分类系统中,肢孔派又被分为绵蟹亚派(Dromiacea)和古短尾亚派(Archaeobrachyura),前者由绵蟹总科(Dromioidea)和人面绵蟹总科(Homolodromioidea)组成,后者由人面蟹总科(Homoloidea)、蛙蟹总科(Raninoidea)和圆关公蟹总科(Cyclodorippoidea)组成[3-5](图1a). 之后,异孔派和胸孔派被合称为真短尾派(Eubrachyura)[6-7](图1b). 但是一些分类学者没有使用肢孔派说法,例如:Martin 和 Davis 将绵蟹亚派提升为派,与真短尾派共同组成短尾下目的两大类群,同时将古短尾亚派中的人面蟹总科归入绵蟹派,蛙蟹总科和圆关公蟹总科归入真短尾派下新设立的蛙蟹亚派(Rainoida)[2](图1c).

图1 短尾下目高阶元的分类系统Fig.1 Classification system of Brachyura
陈惠莲和孙海宝则将绵蟹亚派和古短尾亚派均提升为派,以代替肢孔派,与真短尾派并称短尾下目的三大类群. 该分类系统在《中国海洋生物名录》中得到了沿用[8-9](图1d). 近年来,随着测序技术的发展,分子标记逐渐成为短尾下目分类的重要证据. 部分研究者则提出使用绵蟹派、蛙蟹派(Rainoida)和圆关公蟹派(Cyclodorippoida)、肢孔派[10-11]. 然而,绵蟹派的内部组成依然存在争议,一些研究结果发现人面蟹总科和绵蟹总科形成姐妹群的关系,支持人面蟹总科归入绵蟹派[10,12](图1e);另一些研究结果则显示人面蟹与蛙蟹和圆关公蟹亲缘关系更近,提出应当把人面蟹总科从绵蟹派移入新设立的人面蟹派(Homoloida)之中[13-15](图1f). 然而,在构建不同类群之间的系统进化关系时,分类单元不完全或者分子标记信息不足会导致构建的系统发生树出现长枝吸引的现象,难以准确反映各类群之间自然的进化关系[10,16-17].
与部分基因相比,动物线粒体基因组可以在序列和基因排列顺序两个层面提供丰富、有效的进化信息,已被广泛运用到各类后生动物的系统发生和进化研究中[18-21]. 线粒体基因组全序列数据已经被许多学者们应用于短尾下目的系统发生研究,为解决该类群内部各阶元的系统学争议提供了必要的分子学依据[16,22-25]. 截止目前,已公布的短尾下目的线粒体基因组已超过75种,但科属阶元取样存在不均衡现象,相较于真短尾派,已测定线粒体基因组的物种来自绵蟹总科的仅 1种,人面蟹总科3种,蛙蟹总科3种. 为了增加绵蟹总科不同分类单元的取样,本研究首次选取了绵蟹总科(Dromioidea)绵蟹科(Dromiidae)的典型代表物种——德汉劳绵蟹(Lauridromiadehaani)作为研究对象,应用高通量测序方法测定其线粒体基因组全序列,通过序列比对和比较研究,分析德汉劳绵蟹的线粒体基因组结构和基因排列顺序的进化特征,构建系统发生树,明确德汉劳绵蟹的系统发生位置,解析短尾下目的各派及亚派组成及其之间的系统发生关系.
1 材料与方法
1.1 样本采集和保存
德汉劳绵蟹标本 2017年11 月采集于广西北海,无水乙醇浸泡. 样本鉴定参考《中国海洋蟹类》和《中国动物志 短尾次目 海洋低等蟹类》[8,26].
1.2 DNA提取、扩增及质量检测
取 30 mg 样本组织,浸泡于生理盐水,每 30 min 更换一次生理盐水,脱醇处理. 使用细胞/组织基因组DNA提取试剂盒(generay biotech)进行DNA提取(详细提取步骤参照说明书). 获得的总DNA样品,使用琼脂糖凝胶电泳检测其完整性,使用微量分光光度计检测其浓度及质量. 将质量达标样品送至诺禾致源公司,进行总DNA测序.
1.3 序列组装和基因注释
以毛刺贝绵蟹线粒体基因组(GenBank检索号:KT182070)的13条蛋白编码基因PCGs和2条rRNA基因作为参考序列,使用Geneious 9.1.4 对Illumina 测序获得的原始数据集进行序列组装,得到每个基因相应的重叠序列群. 将相应基因的重叠序列群通过重头组装的方式进行再次组装,获取线粒体基因组全序列. 获得的全序列与毛刺贝绵蟹进行BLAST比对,进行初步的基因注释,使用在线MITOS2对组装序列进行再次识别确认[27].
1.4 线粒体基因组组成与结构特征分析
利用MITOS2在线网址来推测tRNA的二级结构[27]. 使用MEGAX 软件对获得的蛋白编码基因的核苷酸序列进行碱基组成分析. 为了进一步分析绵蟹的线粒体基因组蛋白编码基因的选择压力,利用KaKs_Calculator 2.0对德汉劳绵蟹和毛刺贝绵蟹的13条蛋白编码基因的非同义替换率(Ka)、同义替换率(Ks)及两者的比值(Ka/Ks)进行计算,计算方法使用YN模型[28].
1.5 系统发生分析
从NCBI 数据库中下载24种歪尾类、75种短尾类和28种十足目其他分类单元,共127个物种的线粒体基因组13条蛋白编码基因和2条 rRNA 基因序列,结合德汉劳绵蟹的线粒体基因组序列进行系统发生分析. 将13条蛋白编码基因的氨基酸序列和2条 rRNA 序列分别使用MAFFT 7.215 进行比对[29],使用 GblockV. 0.91b对序列的保守性位点进行选择[30]. 根据比对、修改后的氨基酸序列,将蛋白编码基因的核苷酸序列和 rRNA 序列修改后串联成总的数据集. 按照不同的基因和密码子位将数据集划分成 41 个分区子集,然后使用 PartitionFinder 2确定数据集的最优分区方案和每个分区的最佳核苷酸替换模型,并以此结果修改数据集[31]. 使用IQtree构建最大似然树,ultrafast bootstrap(BS)设置为1000[32-33]. 使用 MrBayes 3.2.6进行贝叶斯分析,蒙特卡洛马尔科夫链(markov chain monte carlo,MCMC)设置运行1 000万代,每1 000代取样一次,其中前 25% 的取样舍弃掉(burn-in)后,余下的取样将被用于汇总出贝叶斯后验概率[34].
2 结果与讨论
2.1 德汉劳绵蟹线粒体基因组结构
德汉劳绵蟹的线粒体基因组是一个环状DNA,全长为15 755 bp,其全部序列被提交至GenBank,检索号为MW239076. 序列分析结果显示其全长序列的碱基组成AT含量71.3%(36.2% A、35.1% T、19.0% C、9.7% G),包含典型的 37 条基因:13 条蛋白编码基因、22 条 tRNA 基因和 2 条 rRNA 基因,其中14条基因在轻链编码,23 条基因在重链编码. 一个主非编码区(main non-coding region,mNCR)长度为566 bp,位于rrnS和trnI基因之间,其AT含量偏高,为80.9%.
和其他短尾下目一样,德汉劳绵蟹线粒体基因组由37条基因紧凑排列而成,13 处相邻基因之间存在碱基对的重叠,重叠长度在 1 bp~7 bp 之间(见表1),这种重叠通常被认为具有防止基因顺序发生重排或防止基因在进化中发生丢失的作用[12,17]. 德汉劳绵蟹线粒体基因组中还存在9处较长的基因间隔(32 bp~130 bp),位于trnE-trnF、trnF-trnT、trnT-nad6、trnS2-trnH、trnH-nad5、nad5-nad4、nad4L-trnP、trnP-nad1和nad1-trnL1的连接处. 已公布的毛刺贝绵蟹同样存在较长的基因间隔(24 bp~482 bp),位于nad5-nad4、trnS2-nad1、nad1-trnL1、trnQ-trnL1、rrnS-trnI和trnI-trnM的连接处[35]. 这些长的间隔序列恰好发生在基因重排的断裂点处,被认为是基因重排时重复的基因拷贝因随机丢失不完全留下的冗余序列,可以作为基因重排的信号为推算基因重排的机制提供有力证据[21].
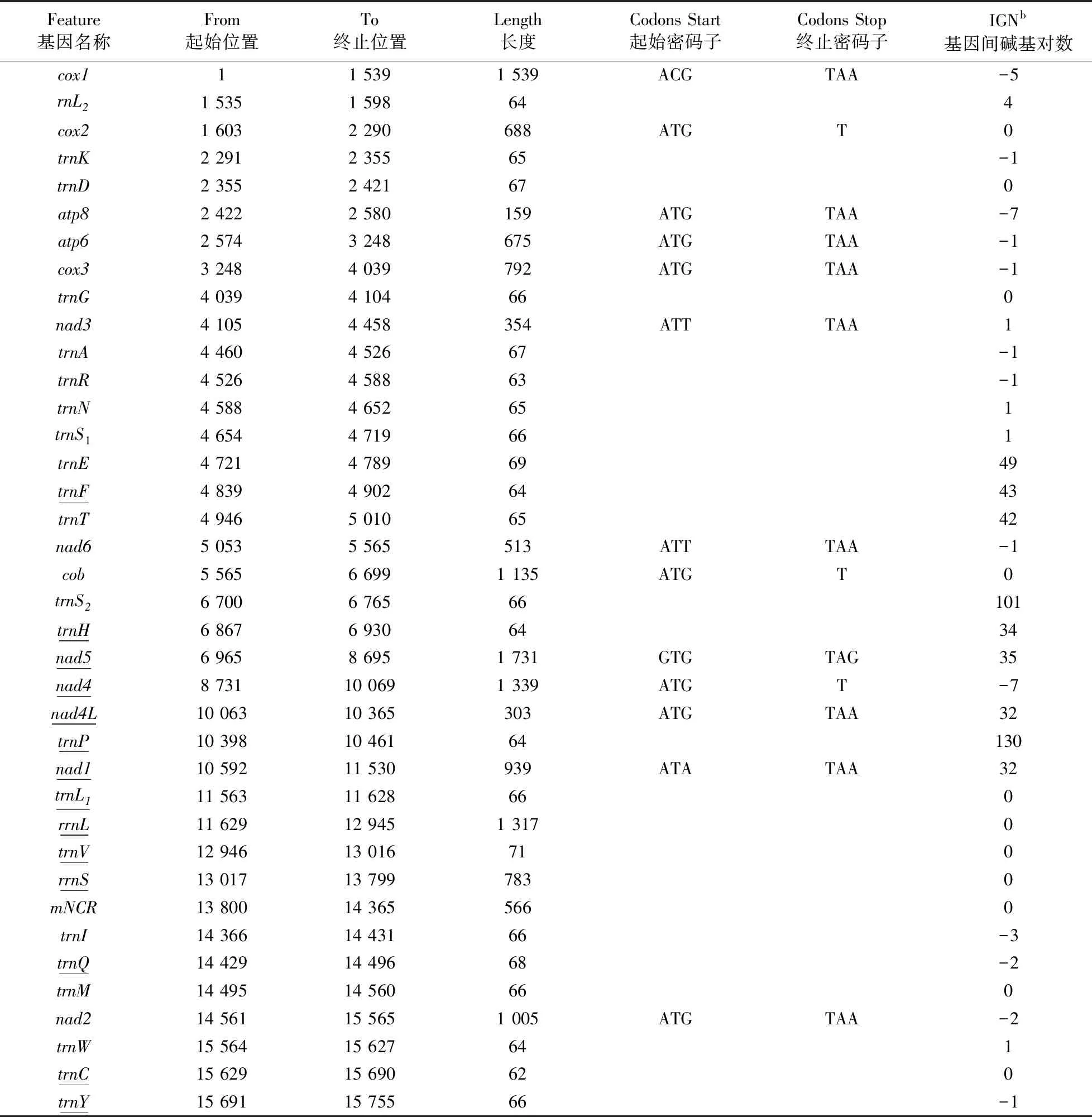
表1 德汉劳绵蟹(Lauridromia dehaani)线粒体基因组特点Table 1 Mitogenomic features of Lauridromia dehaani
2.2 蛋白编码基因
13 条蛋白编码基因中有11条蛋白编码基因的起始密码子为三联体密码子ATN(ATG、ATT、ATA),nad5基因的起始密码子为GTG.cox1基因则以一种不常见的三联体密码子ACG为起始密码子,这种以ACG 作为cox1基因的起始密码子均在蛙蟹科、贝绵蟹科、人面蟹科等低等蟹类的线粒体基因组中发现过,被认为是低等蟹类共同的特征[36]. 蛋白编码基因的终止密码子以TAA、TAG为主,cox2、cob和nad4的终止密码子均为单个碱基T,这种不完整的终止密码子可以通过mRNA成熟过程中多聚腺苷酸化作用补齐、转变为完整的终止密码子[37].
不同呼吸功能的蛋白编码基因的A+T含量和选择压力存在明显的差异. 参与呼吸链的复合体I亚基合成的基因(nad1、nad2、nad3、nad5、nad4和nad6)的A+T含量和Ka/Ks相对较高,分别在70.0%~74.5%和0.033 42~0.065 749;参与复合体 IV亚基合成的基因(cox1、cox2和cox3)和复合体Ⅲ亚基合成的基因(cob)的A+T含量和Ka/Ks值相较于其他基因偏低,分别在64.9%~68% 和0.005 79~0.017 71;参与ATP 合酶合成的atp8基因的AT含量和Ka/Ks最高,说明atp8基因受到的选择压力较小(见图2).

图2 德汉劳绵蟹(Lauridromia dehaani)线粒体基因组13条蛋白编码基因的A+T含量和Ka/Ks值Fig.2 A+T content and Ka/Ks in 13 protein coding genes of Lauridromia dehaani
2.3 tRNA基因
22 条 tRNA基因的长度在62 bp~71 bp,其中trnC最短,trnV最长. 除了trnS1基因缺少 DHU 臂外,其他tRNA基因都具有典型的三叶草二级结构.trnS1基因缺少 DHU 臂的现象在已公布的毛刺贝绵蟹和其他短尾类中也发现过,这种臂的松散或缺失现象被认为和进化压力有关[35-36,38-43].另外,trnS1的反密码子臂由4个碱基对构成,其他tRNA基因均由5个核苷酸对构成.所有tRNA基因的氨基酸接受臂由7个核苷酸对构成. DHU臂和TΨC 臂上分别由2~4 和3~5 个核苷酸对构成. 22 条tRNA的接受臂、DHU臂、反密码子臂和TΨC臂上出现G/U的错配24次,trnY的反密码子臂上出现U/U错配,trnK的接受臂上出现 C/U 错配(见图3).
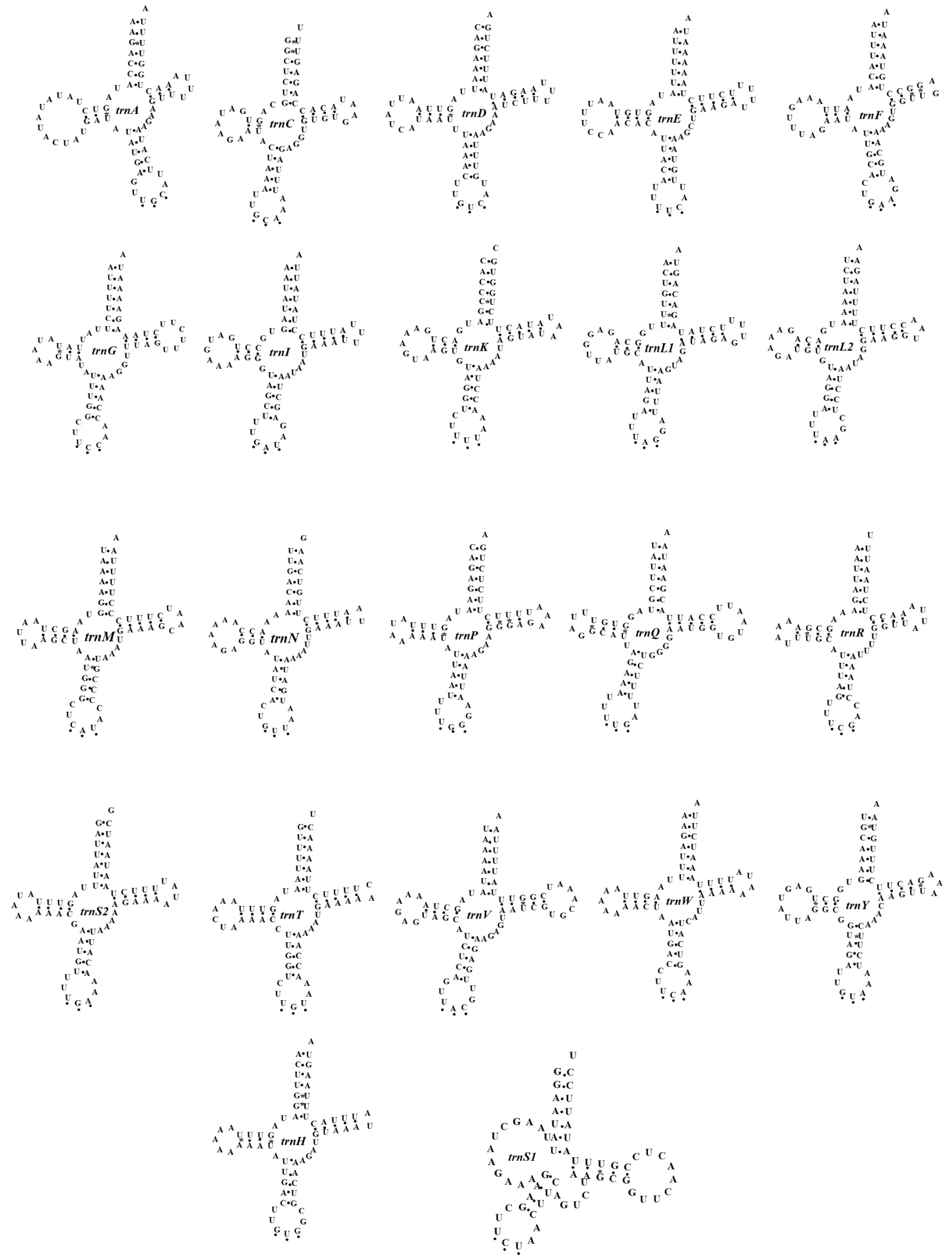
图3 德汉劳绵蟹(Lauridromia dehaani)22条tRNA二级结构Fig.3 22 tRNA secondary structures of Lauridromia dehaani
2.4 基因重排
与短尾类线粒体基因组原始的基因排列比较[21],德汉劳绵蟹的线粒体基因组发生了显著重排:基因块[nad6-cob-trnS2]从trnP和nad1基因之间易位到trnF和nad5基因之间;trnT基因从nad4L和trnP基因间易位到基因块[nad6-cob-trnS2]的上游;trnF易位到trnE和trnT之间,最终在trnE与nad1基因之间形成新的排列顺序,即[trnF-trnT-nad6-cob-trnS2-trnH-nad5-nad4-nad4L-trnP].毛刺贝绵蟹仅trnH、trnQ和mNCR发生了易位:trnQ从trnI和trnM基因之间易位到trnL1和rrnL基因之间,mNCR从rrnS和trnI基因之间易位到trnL1和trnQ基因之间[35](图4). 根据断裂点和串联重复随机丢失模型[20-21],推断出德汉劳绵蟹的线粒体基因组重排的途径(图4):首先基因片段[trnH-trnF-nad5-nad4-nad4L-trnT-trnP-nad6-cob-trnS2]发生一次重复,然后第一个片段中trnH、[nad5-nad4-nad4L]和trnP丢失,第二个片段中trnF、trnT和[nad6-cob-trnS2]丢失,由于重复基因不完全丢失会在trnE-trnF、trnF-trnT、trnT-nad6、trnH-nad5、nad4L-trnP和trnP-nad1的基因连接处留下较长的基因间隔(32 bp~130 bp). 另外,trnS2-trnH和nad5-nad4的基因连接处的间隔是由于trnH从[nad5-nad4]基因块易位到基因块[nad3-nad5]之间时留下.师国慧等人曾对毛刺贝绵蟹线粒体基因发生重排的路径进行了推导:首先基因块[rrnL-trnV-rrnS-mNCR-trnI-trnQ]经历一次串联重复,随后丢失了第一个拷贝中的[rrnL-trnV-rrnS]和trnI,第二个拷贝中的mNCR和trnQ,最终形成毛刺贝绵蟹线粒体基因排列顺序.依据此推断结果,nad1和trnL1基因之间不会存在长的基因间隔,但是实际发现两个基因之间存在482 bp 的基因间隔,因此推断重复的起始位置为trnL1,并且基因块[trnL1-rrnL-trnV-rrnS-mNCR-trnI-trnQ]经历两次串联重复,随后第一个拷贝片段上的基因全部丢失,最终在nad1和trnL1基因之间留下长片段的非编码区.另外,在毛刺贝绵蟹线粒体基因组中,trnQ-trnL1、rrnS-trnI和trnI-trnM处的基因间隔经过 MITOS2 重新被确认为 137 bp、270 bp 和31 bp.
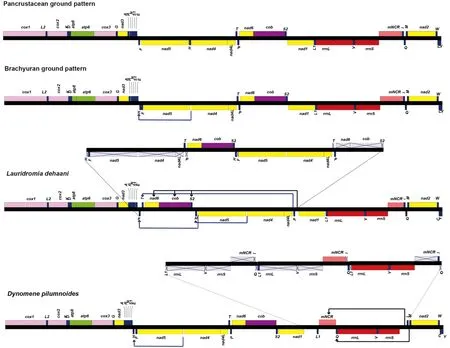
图4 德汉劳绵蟹(Lauridromia dehaani)与毛刺贝绵蟹(Dynomene pilumnoides)线粒体基因组排列顺序的比较及其可能的重排机制Fig.4 Comparisons of mitochondrial gene orders and putative rearrangement process for Lauridromia dehaani and Dynomene pilumnoides
基因排列可以作为特定谱系和分类群的共衍生性状,为系统发生重建和进化关系推导提供重要的分子证据[18-25]. 已有研究发现,trnH从[nad5-nad4]基因块易位到基因块[nad3-nad5]之间被认为是短尾类线粒体基因组所共享的特征[21],并被证实在已测定的短尾类线粒体基因组中均存在[23,25]. 本研究中德汉劳绵蟹和已公布的毛刺贝绵蟹均存在trnH易位现象,提示绵蟹应归置于短尾下目. 与短尾类线粒体基因组原始排列顺序相比[25],德汉劳绵蟹和毛刺贝绵蟹的线粒体基因组基因排列顺序存在着明显差异,前者发生巨大的基因重排,涉及到nad6、cob、trnH、trnF、trnS2和trnT等6条基因发生了易位,后者仅trnQ和mNCR2条基因发生了易位,这两种独特的排列顺序是否在绵蟹科和贝绵蟹科的其他类群中也存在,还需今后获得更多的样本加以验证.
2.5 短尾下目的各派及亚派的系统发生关系
基于 13 条蛋白编码基因和 2 条rRNA基因联合数据集构建的最大似然树(ML)和贝叶斯树(BI)得到了几乎一致的拓扑结构(图5所示). 结果显示,24种歪尾类物种和75种短尾类物种各聚成一支,支持度均为 BS/BPP=100/1.00. 短尾下目的各派及亚派之间展现出强健的拓扑结构:(绵蟹总科,人面蟹总科),(蛙蟹总科,真短尾派),其中德汉劳绵蟹和毛刺贝绵蟹聚成的一支(BS/BPP=100/1.00),再与人面蟹总科的物种形成姐妹群关系,成为短尾下目的基部分支,支持度均为 BS/BPP=100/1.00;蛙蟹总科的3个物种聚成一支,与胸孔亚派和异孔亚派组成的真短尾派形成姐妹群关系,支持度为 BS/BPP=98.9/1.00;在真短尾派中,来自异孔亚派的溪蟹科(Potamidae)和束腰蟹科(Parathelphusidae)的物种聚成一个分支,并与胸孔亚派的类群形成姐妹群的关系,节点支持度为BS/BPP=100/1.00.
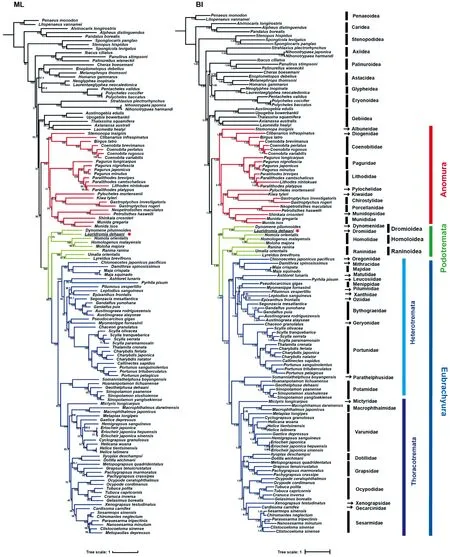
图5 基于13 条蛋白编码基因(PCGs)和2条rRNA基因的构建的最大似然树(ML)和贝叶斯树(BI)自举值(BS)和贝叶斯后验概率(BPP)均在内部节点显示Fig.5 Phylogenetic trees derived for Brachyura using maximum likelihood(ML)and Bayesian inference(BI)analyses based on 13 protein coding genes(PCGs)and two rRNAs. Maximum likelihood bootstrap(BS) and Bayesian posterior probabilities(BPP)values were shown at the corresponding nodes
在所有蟹类中,绵蟹的成体还有退化的尾肢,鳃数甚多,有14~20对丝状鳃(非叶状鳃)[8];从个体发育来看,蚤状幼体没有长的背刺,第3颚足无外肢,具有歪尾类(Anomura)的特征,因此早期有些学者将其归入歪尾下目[44-46],该结果得到了Speares 使用18S rRNA 作为分子标记的支持,但是节点支持度薄弱[47]. 截止目前,大部分的分类学者认为绵蟹的成体更接近真正蟹类的外形特征,即其头胸部发达,腹部极为退化并折附于头胸部腹面,而后大量的分子系统发生树也显示绵蟹和和其他短尾类具有紧密的亲缘关系,均支持将绵蟹归入短尾下目[7-11,26,48-49]. 本研究中,联合分子系统发生关系和线粒体排列顺序的共衍性状共同佐证绵蟹派隶属短尾下目.
研究结果还发现,如果使用肢孔派的分类观点,肢孔派的绵蟹总科、人面绵蟹总科、人面蟹总科、蛙蟹总科和圆关公蟹总科应该聚为一支,然而本研究中蛙蟹总科与真短尾派形成姐妹群的关系,不支持肢孔派的单系性. 越来越多的分子和形态证据支持使用多派分类的方法以替代肢孔派. 然而纵观各学派的分类观点时,各派内部的组成和系统发生关系没有得到统一,特别是人面蟹总科的系统发生地位摇摆不定:或与圆关公蟹总科和蛙蟹总科亲缘关系近支持三者组成古短尾派[8-9](图6a);或与(绵蟹总科+人面绵蟹总科)形成姐妹群关系支持人面蟹总科归置于绵蟹派[2,10,48,50],同时圆关公蟹总科和蛙蟹总科形成姐妹群关系(图6b);或与(圆关公蟹总科+蛙蟹总科+真短尾派)的分支形成姐妹群关系支持单独设立一派,但圆关公蟹总科和蛙蟹总科哪个与真短尾派亲缘关系更近也未得到一致结论[13,15,51](图6c、d). 相较于Shi等人使用线粒体基因组蛋白编码基因构建的短尾下目的系统发生树[36],本研究增加了绵蟹总科的取样避免长枝吸引对树结构的影响[16]. 研究结果显示,人面蟹总科与绵蟹总科,蛙蟹总科与真短尾派具有紧密的亲缘关系,节点支持度明显增加. 遗憾的是,由于缺少圆关公蟹总科和人面绵蟹总科的样本,这5个总科之间的系统发生关系依然比较模糊,亟需获取其他两个总科的代表物种的线粒体基因组全序列构建系统发生树加以解析.

图6 短尾下目内部个派之间的系统发生关系的假说Fig.6 Hypotheses of interrelationships among brachyuran section
在真短尾派中,若按照雌、雄生殖孔分别在第六胸节及第四步足底节的特征,原生淡水蟹应被分配到异孔亚派[7]. 然而,Von Sternberg和Cumberlidge 根据更为细致的雄性生殖孔的解剖特征,提出溪蟹科、陆溪蟹科(Gecarcinucidae)和拟束腹蟹科(Pseudothelphusidae)应放置在胸孔亚派[52-53]. 本研究结果显示溪蟹科(Potamidae)和束腰蟹科(Parathelphusidae)的代表物种聚成的分支与胸孔亚派形成姐妹群的关系,且节点支持度高,证实了之前使用部分基因、线粒体基因组和转录组序列推断的系统发生结果,支持溪蟹科和束腰蟹科与胸孔亚派的亲缘关系更近[11,16,22,54]. 但与Tsang等人综合部分线粒体基因和核基因的重建的系统发生关系相矛盾,其研究结果显示旧大陆的非洲溪蟹科(Potamonautidae)、溪蟹科和陆溪蟹科的代表物种形成的一个单系类群,成为异孔亚派的早期分支,但是节点支持度薄弱[10]. 虽然线粒体基因组序列能够构建出强健的系统发生树,若要进一步厘定淡水蟹的分类地位以及追溯淡水蟹的起源,仍需要针对淡水蟹类、胸孔亚派和其他异孔亚派等类群进行广泛取样,重建它们之间的系统发生关系.
3 结论
本研究首次测定德汉劳绵蟹的线粒体基因组,确定其序列全长为15 755 bp,其蛋白编码基因和 tRNA 呈现以下进化特征:不同呼吸功能的蛋白编码基因的碱基组成(AT含量)和选择压力(Ka/Ks)呈现差异,atp8基因最高,cox1基因最低;cox1以低等蟹类中常见的 ACG 作为起始密码子;trnS1缺失了DHU臂. 绵蟹总科的德汉劳绵蟹线与该总科的毛刺贝绵蟹的线粒体基因组排列顺序完全不同,意味着该总科其他类群存在基因重排的潜能,两者又共享短尾类线粒体基因组中trnH易位特征,为支持绵蟹总科归置于短尾下目提供了独立佐证. 系统发生树解析出强健的拓扑结构,揭示出绵蟹总科与人面蟹总科之间,蛙蟹总科与真短尾派之间均形成姐妹群关系,为短尾下目高阶元分类提供分子依据.
