不同带电特性的壳聚糖/酪蛋白相互作用研究
2021-03-06傅玉颖沈亚丽陈国文
张 超,傅玉颖,2*,沈亚丽,陈国文,2,张 豪,靳 冰
(1 浙江工商大学食品与生物工程学院 杭州310018 2 浙江工商大学杭州商学院 杭州310018)
多糖/蛋白质作为食品结构的基础物质,对食品的物理、化学性质起决定性的作用。现代食品制造除以营养学为基本外,还兼顾质构设计。近些年,越来越多的研究者开始着力于从底层结构设计食品,从而实现对食品质构的准确调控。例如:设计不同质构的凝胶体系以及不同的蛋白质-多糖复合物体系等。多糖/蛋白质因相异的电性和易于复合等性质而得到关注。利用蛋白质-多糖复合后物理、化学性质的不同可构建各异的应用场景,例如用酪蛋白酸钠-可溶性淀粉复合物可以制备可食性薄膜[1],乳蛋白/黄原胶复合后可以充当脂肪或肉类的替代品等[2]。
多糖、蛋白质在复合过程中受到内、外环境的影响而形成具有不同微结构的产物,二者间相互作用的最终产物往往具有各异的宏观性质,例如不同的流动特性及黏弹特性等。多糖/蛋白质间的相互作用可以从外部因素(pH 值、离子强度、温度等)调控。pH 值通过改变多糖和蛋白质的带电性及带电量,影响二者间的静电相互作用;离子强度如高浓度的Na+会屏蔽两者间的电荷效应[3],使得体系变得不稳定;温度的升高则会导致蛋白质变性,结构舒展,使内部的带电基团大量暴露,与多糖的凝聚反应强度增加[4]。从内部因素(分子质量、结构等)看,多糖分子质量的增加,促进了多糖与蛋白质的结合,从而有利于形成静电复合物[5]。Bekale 等[6]讨论了壳聚糖(CS)与牛和人血清蛋白间的相互作用,得出蛋白质二级结构的差异,最终导致CS 与两种模式蛋白间相互作用力的不同。以上几种因素的影响已有大量的研究,而基于多糖本身的带电特性对多糖/蛋白质复合的影响研究较少。利用大分子的带电特性来调控多糖与蛋白质的非共价相互作用十分关键。
CS 目前作为自然界中唯一的阳离子多糖,具有天然无毒,良好的生物相容性,可降解等优点[7],因此在食品、化妆品、生物医学和制药工程领域的应用前景被普遍看好。壳聚糖脱乙酰度(D.D)的差异影响CS 带电特性。如王伟等[8]在研究CS的Mark-Houwink 方程时,发现D.D 较高的CS 氨基质子化后形成的阳离子聚电解质分子链段间以及链段与溶剂间的相互作用增强,有效电荷密度增加使得聚电解质溶液黏度增大。酪蛋白又称干酪素,是牛乳中特有的一组含有大量钙、磷的蛋白质[9]。正是由于酪蛋白独特的胶束结构,其与多糖的复合物可被用作脂肪替代品以及肉模拟物[10]。酪蛋白分子的可降解性使其稳定性较差[11],而CS被广泛用于蛋白质的稳定剂,提高了蛋白质的活性与稳定性[12]。CS 与酪蛋白通过非共价相互作用复合,使复合物具有更好的稳定性和功能性。
本试验选用不同带电特性的CS 和酪蛋白为研究对象,采用荧光光谱、紫外-可见吸收光谱、傅里叶红外吸收光谱及黏度分析法研究CS 与酪蛋白的复合机制及流变特性。
1 材料与方法
1.1 试验材料
壳聚糖分子质量约740 ku,南通兴成生物制品厂;酪蛋白(Lot#SLBL6515V),美国Sigma-Aldrich 公司;氢氧化钠、盐酸,杭州市化学试剂有限公司;氯化钠、乙酸(冰乙酸)、无水乙酸钠,上海麦克林生化科技有限公司;氢氧化钠、盐酸、氯化钠、乙酸(冰乙酸)、无水乙酸钠均为分析纯试剂。
1.2 试验仪器与设备
pH 计(PHS-3C),上海理达仪器厂;激光粒度分析仪(Zeta sizer Nano ZS),英国马尔文公司;荧光分光光度(RF-5301PC),日本岛津公司;紫外-可见分光光度计(UV-2600),日本岛津公司;傅里叶变换红外光谱仪(NicoletiS5),上海力晶科学仪器有限公司;旋转流变仪(MCR302),奥地利安东帕有限公司。
1.3 试验方法
1.3.1 样品溶液的制备 CS 和酪蛋白储备溶液:准确称取一定质量的CS 溶于0.1 mg/mL 乙酸溶液中,于1 000 r/min 的磁力搅拌器上匀速搅拌3 h,置于4 ℃冰箱贮藏过夜,保证充分水合,制得质量浓度为8 mg/mL 的CS 溶液。称取一定质量的酪蛋白溶于去离子水中,在磁力搅拌器上搅拌溶解,配成质量浓度为2 mg/mL 的酪蛋白溶液。
1.3.2 测定CS 脱乙酰度 准确称取一定质量的CS 样品于烧杯中,用0.1 mol/L 盐酸在室温下充分溶解。将pH 计电极置于溶液中,用0.1 mol/L NaOH 溶液滴定,记录消耗NaOH 的体积及相应的pH 值,绘制pH 值与消耗NaOH 体积的关系曲线以及CS 的双突跃滴定曲线。

式中:c——消耗NaOH 浓度(mol/L);V2——第1 次pH 值突变时消耗的NaOH 体积(mL);V1——第2 次pH 值突变时消耗的NaOH 体积(mL);m——CS 样品的质量(g)。
1.3.3 测定不同脱乙酰度CS 带电特性 一般情况下,研究者利用Zeta 电位测量多糖/蛋白质复合后粒子的带电特性。从微观角度看,带电的分散粒子在溶液中相互靠近时形成扩散双电层。当扩散层内的粒子相对于层外粒子发生迁移时会形成滑动面,在滑动面测得的电荷即粒子的表面电荷б[13]。其中,Zeta 电位中粒子的迁移速率与表面电荷б 密切相关。而多肽链作为蛋白质的基础构成部分,与多糖复合后多肽链极性改变,链段伸展或收缩,表面电荷相对不稳定。为了表征粒子的真实电荷,采用Ohshima 软粒子模型,Ohshima 软粒子的迁移率不取决于表面电荷б,而取决于体积电荷密度ZN[14]。在复合体系中,粒子体积的变化导致粒子结构、电子态密度以及电荷分布的改变。Ohshima 软粒子模型可以反映粒子在聚合体系中的真实电荷。
在25 ℃条件下分别测定0.1% CS 在10,40,60,80 mmol/L NaCl 溶液的电泳迁移率(μE)。以μE对NaCl 溶液的浓度绘制曲线图,用公式拟合CS在区域空间的电荷密度N 与代表这些电荷价态的Z,以及代表在聚电解质(CS)区域阻止液体流体的程度。其公式如下:

式中:μ——CS 的电泳淌度(m2/s·V);ε0——CS 溶液的真空绝对介电常数;εr——CS 溶液的相对介电常数;η——介质黏度(Pa·s);ψ0——表面区域和溶液的边界的电势(V);Km——表面区域的Debye-Hückel 有效参数;k——玻尔兹曼常数;T——绝对温度(K);z——溶液离子价态(NaCl 溶液中z=1);n——CS 溶液的浓度(mol/L);e——基本电荷(C);ψDON——Donnan 电势(V);N——电解质区域电荷密度(C/m3);Z——相应的电荷价态。
1.3.4 荧光光谱 将酪蛋白与CS 分别置于303 K 和313 K 恒温水浴中30 min,采用荧光分光光度对pH 6.0 的酪蛋白和CS-酪蛋白溶液在波长290~500 nm 范围扫描,设定荧光激发和发射狭缝宽度均为5.0 nm,采样间隔1.0 nm,激发波长280 nm,记录溶液在336 nm 附近荧光强度的变化。
1.3.4.1 荧光猝灭机制 荧光物质激发态分子的寿命和猝灭剂的浓度直接影响动态猝灭的效率,
其过程遵循Stern-Volmer 方程[15]。

式中:F0——猝灭剂加入前(CS)的荧光强度(A.U.);F——猝灭剂加入后(CS-酪蛋白)的荧光强度(A.U.);[Q]——猝灭剂(CS)的浓度(mol/L);Kq——猝灭速率常数;Ksv——Stem-Volmer 猝灭常数;τ0——分子聚合物的荧光平均寿命(10-8s)[16]。在动态猝灭中,猝灭剂对分子聚合物的最大扩散碰撞速率常数为2.0×1010L/(mol·s)[17]。
1.3.4.2 结合位点数和结合常数 假设酪蛋白分子上对CS 有n 个等同的且独立的结合部位,结合常数为KA,则CS-酪蛋白间的相互作用过程中,结合常数KA 和结合位点数n 符合下面Lineweaver-Burk 双倒数公式[18]。

式中:F0——猝灭剂加入前(CS)的荧光强度(A.U.);F——猝灭剂加入后(CS-酪蛋白)的荧光强度(A.U.);[Q]——CS 的浓度(mol/L);KA——结合常数;n——结合位点数。
1.3.4.3 作用力类型 多糖/蛋白质间的非共价相互作用主要是由静电相互作用、疏水效应、氢键及范德华力等驱动[19]。Ross 等[20]根据热力学常数(焓变值ΔH、熵变值ΔS)的符号和大小总结出高分子聚合物与其它物质之间结合的热力学规律:ΔH、ΔS 均为正值,两者间的相互作用力主要为疏水作用力;ΔH、ΔS 均为负值,两者间的相互作用主要是范德华力或氢键;ΔH<0,ΔS>0,两者间的相互作用主要是静电相互作用。
在测量的温度范围变化不大时,ΔH、ΔS 可看作常数,由Vant Hoff 等式和热力学公式可得CS-酪蛋白间的热力学参数[21]。

式中:K1,K2为303 K 和313 K 温度下的结合常数;R——气体常数;T——热力学温度(K)。
1.3.5 紫外-可见吸收光谱 在25 ℃条件下,采用紫外-可见分光光度计测定各组溶液在pH 6.0时的吸光值。波长范围200~800 nm,扫描速度为中速,采样间隔1 nm,扫描方式为自动,狭缝宽度0.2 mm,记录紫外吸收值的变化。
1.3.6 傅里叶变换红外光谱(FTIR)称取一定质量的CS、酪蛋白及冷冻干燥后的CS-酪蛋白复合物(pH 7.5)样品,按质量比1∶100 的比例与KBr混合、研磨,用压片机将研磨均匀的粉末制成透明薄片。在25 ℃下,用傅里叶变换红外光谱仪在波数400~4 000 cm-1范围,以4 cm-1的分辨率扫描样品16 次。
1.3.7 流变稳定黏度测定 利用旋转流变仪测定酪蛋白和CS-酪蛋白复合溶液的表观黏度,选取CC27 型号的同心圆筒模式转子。温度设置为25℃,剪切速率范围0.01~100 s-1,记录样品表观黏度随剪切速率的变化。
Power-law 模型常被用来描述复合溶液的流变行为。

式中,б——剪切应力(mPa);κ——剪切速率(S-1);γ——稠度系数(Pa·sn);κ 值与复合溶液的黏稠度成正比。n=1,理想牛顿流体,表观黏度不随剪切速率而变化;n>1,胀塑性流体,即剪切增稠;n<1,假塑性流体,具有剪切变稀的特征,n 越小表示假塑性程度越大[22]。
1.3.8 数据分析 本试验中的结果平均重复3次,数据用Excel 2010 与SPSS 24.0 软件统计分析,P<0.05 表示显著,P>0.05 表示不显著。用Origin 2017 软件作图。
2 结果
2.1 CS 脱乙酰度
由图1 可知,在滴加NaOH 时,整个体系中出现两个突变点。第1 次突变是由于NaOH 中和溶液中的HCl,第2 次突变是NaOH 与CS 的氨基反应的结果。由公式(1)计算得到本试验中测得的CS 的D.D 值分别是82.5%和94.6%。为了便于表述,将两种CS 简称为LDCS 与HDCS。
2.2 不同脱乙酰度CS 带电特性
由图2 可知,μE与NaCl 溶液的浓度呈负相关。表1 中,D.D 为82.5%和94.6%的CS 的ZN 值分别是0.0077 mol/L 和0.0110 mol/L,表明随着CS脱乙酰度的增加,ZN 也随着升高。这是由于CS 分子链上的氨基质子化,使得CS 在溶液中的带电基团增加,带电量提高。表2 中,拟合得到的软粒子电位几乎是所测电位的一半,可能是因为软粒子模型中带电粒子极化,对可穿透层内的粒子起到屏蔽作用[23]。
2.3 CS-酪蛋白复合溶液的荧光光谱分析
2.3.1 荧光猝灭机制分析 图3 是不同脱乙酰度的CS 对酪蛋白的荧光猝灭光谱图。当激发波长为280 nm 时,酪蛋白在波长336 nm 附近出现最大荧光发射峰,其主要由酪蛋白中色氨酸(Trp)残基产生。在波长336 nm 处,CS 的加入,使得酪蛋白的荧光强度逐渐降低,表明CS 与酪蛋白发生分子内或分子间的相互作用,这也说明CS 对酪蛋白有一定的荧光猝灭效果。同时,添加同浓度的CS,HDCS 对酪蛋白的荧光猝灭效果更明显。这主要是由于HDCS 分子链上的带电基团增加,与酪蛋白分子结合后折叠缠绕,比LDCS 对Trp 残基覆盖全面,因此猝灭效果更明显。

图2 不同脱乙酰度CS 的电泳迁移率Fig.2 Electrophoretic mobility of chitosan with different degrees of deacetylation

表1 不同D.D 值的CS 的电荷密度ZNTable 1 Charge density ZN of chitosan with different degrees of deacetylation

表2 不同D.D 值CS 的电位与基于Ohshima 软粒子模型电位比较Table 2 Comparison of ζ-potentials of chitosan with different degrees of deacetylation and potentials based on Ohshima soft particle model
如图4所示,以CS 的浓度[Q]为横坐标,F0/F为纵坐标,制得温度303K 和313K 时不同D.D 的CS 猝灭酪蛋白的Stern-Volmer 图,斜率即Ksv。表3 是根据公式8 的得到的Stern-Volmer 常数(Ksv)和双分子速率猝灭常数(Kq)。在温度303 K和313 K 时的双分子速率猝灭常数值均大于最大扩散碰撞速率常数2.0×1010,这表明CS 对酪蛋白的猝灭方式为静态猝灭。Stern-Volmer 常数(Ksv)与温度呈反比,即Stern-Volmer 常数(Ksv)随着温度的升高而下降,这说明CS 对酪蛋白的猝灭方式是静态而非动态的。

图3 CS 对酪蛋白的荧光猝灭光谱图Fig.3 The quenching fluorescence spectrometry of chitosan on casein intensity

图4 不同温度下CS 猝灭酪蛋白的Stem-Volmer 图Fig.4 Stem-Volmer curve of chitosan quenching casein at different temperatures

表3 不同温度下CS-酪蛋白相互作用的Stern-Volmer 常数和双分子速率猝灭常数Table 3 Stern-Volmer constant and bimolecular rate quenching constant of CS-casein interaction
2.3.2 结合位点数和结合常数的确定 如图5所示,在不同温度下,以lg[Q]为横坐标,lg[(F0-F)/F]为纵坐标作图,直线的斜率和截距分别代表结合位点数n 和结合常数KA。计算得出的结合位点数n 和结合常数KA 见表4。HDCS 在303K 和313K时的结合位点数分别为1.29 和1.36,在相同条件下LDCS 的结合位点数分别是1.47 和1.44。二者的结合位点n 均在1 附近,说明不同D.D 的CS 与酪蛋白均只有一个结合位点。
2.3.3 作用力类型的分类 由公式(10)、(11)、(12)求得CS-酪蛋白间相互作用的ΔH、ΔS 和吉布斯自由能ΔG,结果见表5。
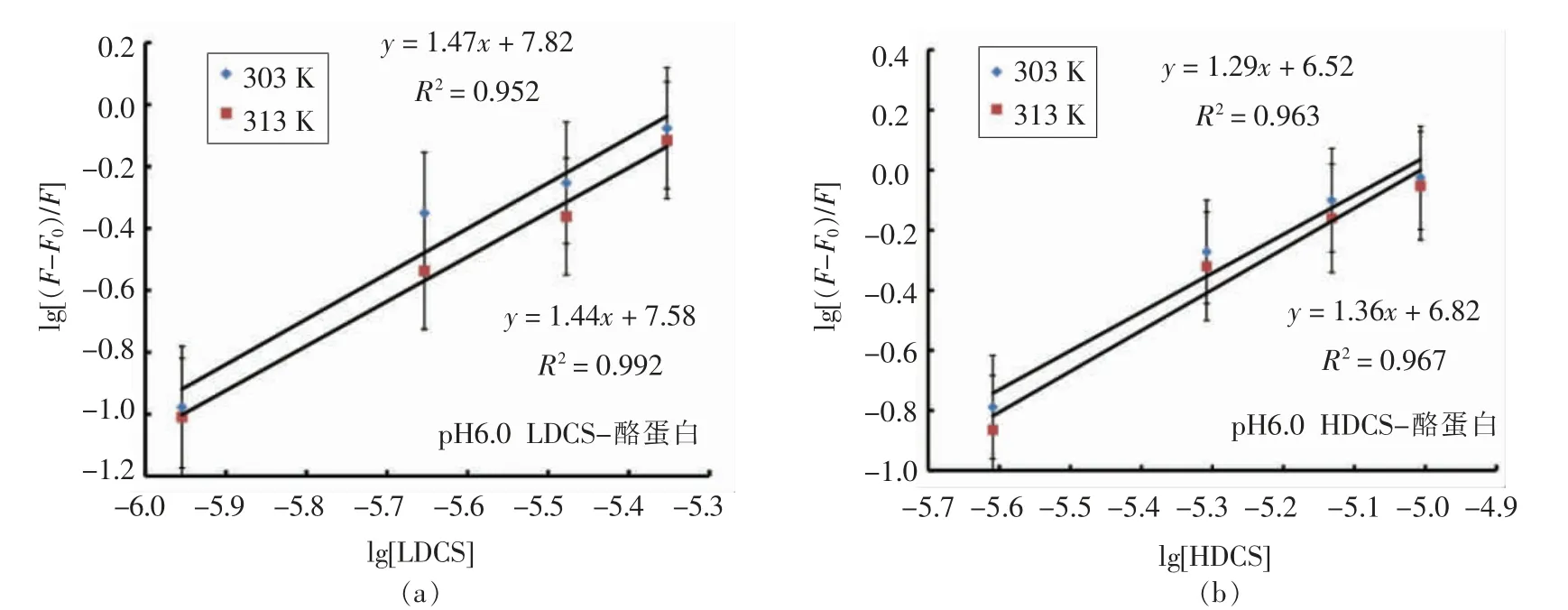
图5 不同温度下CS 猝灭酪蛋白的双对数图Fig.5 Double logarithmic plot of CS quenching protein at different temperatures

表4 不同温度下CS 和酪蛋白的结合常数和结合位点数Table 4 Binding constant and binding site number of CS and casein at different temperatures

表5 CS 和酪蛋白相互作用的热力学参数和主要作用力类型Table 5 Thermodynamic parameters and major types of interaction between CS and casein
表5 中,在pH 6.0,温度303K 和313K 条件下的吉布斯自由能△G<0,表明CS 和酪蛋白间的相互作用是自发的。HDCS-酪蛋白间的相互作用过程中△H<0,△S>0,表明二者的相互作用力以静电力作用为主,CS 和酪蛋白复合过程中,CS 的正电荷与酪蛋白中的负电荷发生静电力相互作用。而LDCS-酪蛋白间的相互作用过程中△H>0,△S>0,表明二者间主要以疏水作用力为主,这可能是由于pH 6.0 时LDCS 分子结构中的疏水基团多于HDCS。
2.4 CS-酪蛋白复合溶液的紫外-可见吸收光谱分析
如图6所示,在pH 6.0 时,酪蛋白溶液紫外特征吸收峰在波长275 nm 附近,主要是由酪氨酸(Tyr)残基贡献。随着CS 的加入,酪蛋白的紫外吸收峰值逐渐增加,这说明CS 氨基与酪蛋白之间发生分子间或分子内的相互作用。添加LDCS 的酪蛋白溶液紫外吸收峰基本不变,说明LDCS 的加入未使酪蛋白中Tyr 的微环境发生改变。相反的,添加HDCS 的酪蛋白复合溶液紫外最大吸收峰出现轻微红移(像右移动),表明HDCS 的加入使得酪蛋白中的Tyr 的微环境发生改变,导致其极性升高,疏水性降低。
2.5 CS-酪蛋白复合物的傅里叶红外光谱(FTIR)分析
图7 中,从CS、酪蛋白和CS-酪蛋白复合物中都检测到-OH 伸缩振动吸收峰,分别在3 440,3 436,3 434 cm-1处。酪蛋白的FTIR 图谱显示,1 637 cm-1处的吸收峰是酰胺Ⅰ带-C=O 的伸缩振动,1 540 cm-1的吸收峰是酰胺II 带-C-N 的伸缩振动和-N-H 的弯曲振动,1 439 cm-1为-C-H 的弯曲振动。CS 的FTIR 图谱中,在1 659 cm-1处的吸收峰是酰胺Ⅰ带-C=O 的伸缩振动,1 596 cm-1处的吸收峰是-N-H 的弯曲振动,1 420 cm-1处是-CH2的弯曲振动,1 375 cm-1处是-C-H 的伸缩运动,1 319 cm-1处是酰胺Ⅲ带-C-NH2的伸缩振动,1 155 cm-1处是C-O-C 的对称收缩,1 081 cm-1处的吸收峰是-C-O 的拉伸振动;CS-酪蛋白复合物的FTIR 光谱图中出现1 569 cm-1和1 652 cm-1两个新的吸收峰,CS 在1 596 cm-1处的吸收峰与酪蛋白在1 637 cm-1吸收峰的消失,说明CS 中的-NH3+基团和酪蛋白中-COO-基团发生相互作用。
2.6 CS-酪蛋白复合物的流变稳定黏度分析
如图8所示,随着剪切速率的增加,CS-酪蛋白复合物流变稳态黏度总体呈现下降趋势,且HDCS-酪蛋白在整个剪切速率范围的黏度要高于LDCS-酪蛋白。如表6所示,HDCS-酪蛋白复合体系的幂律方程中κ 明显大于LDCS-酪蛋白,正好验证了图8 的结论。此外,复合体系的n 均小于1,表明该复合体系为非牛顿流体,在较高的剪切速率下,整个体系的黏度趋于稳定,出现剪切变稀的现象,说明CS-酪蛋白复合体为典型的假塑性流体。

图6 CS-酪蛋白复合溶液的紫外-可见吸收光谱图Fig.6 UV-visible absorption spectrum of CS-casein complex solution

图7 CS、酪蛋白及CS-酪蛋白复合物的FTIR 光谱图Fig.7 FTIR spectra of CS,casein and CS-casein complex

表6 CS-酪蛋白复合体系的幂律方程参数Table 6 Power law equation parameters of CS-casein complex system
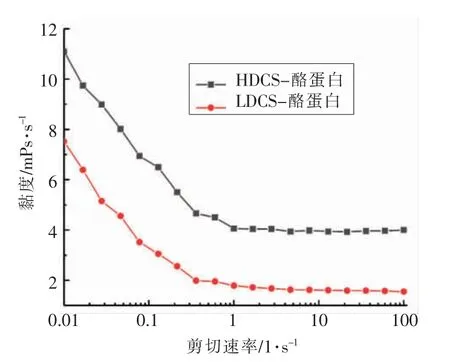
图8 CS-酪蛋白复合物流变稳态黏度随剪切速率的变化曲线Fig.8 Curve of steady-state viscosity versus shear rate of CS-casein complex
3 结论
基于多糖本身的带电特性,研究了不同带电特性的CS 与酪蛋白的非共价相互作用。通过荧光光谱、紫外-可见光谱、红外吸收光谱等表征方法弄清CS 中的-NH3+基团和酪蛋白中-COO-基团发生了相互作用。HDCS-酪蛋白间的相互作用主要为静电相互作用,LDCS-酪蛋白则主要为疏水相互作用。流变稳态黏度分析显示,HDCS-酪蛋白复合物的黏度要高于LDCS-酪蛋白复合物,说明CS-酪蛋白复合体为典型的假塑性流体。
