维氏气单胞菌中AHLs介导的群体感应现象及大蒜提取物的干扰作用
2021-03-06赵丹丹丁玉庭陈文烜周绪霞
赵丹丹,丁玉庭,陈文烜,胡 俊,周绪霞*
(1 浙江省农业科学院食品科学研究所 杭州310021 2 浙江工业大学食品科学与工程学院 杭州310014)
微生物的生长繁殖和代谢是导致食品腐败变质的重要因素。细菌群体感应(Quorum sensing,QS)是一种依赖于细胞密度的信息交流方式,以自诱导物(Autoinducers,AIs)为群体感应信号分子,它通过调控食品中优势腐败菌的生长代谢来调控食品腐败进程[1]。研究表明,群体感应抑制剂可通过与AIs 受体蛋白结合,降解AIs 分子和阻断群体感应通路等途径有效干扰细菌群体感应的表达,降低食源性细菌的致病性或致腐性[2]。利用群体感应抑制剂靶向调控细菌的群体感应系统来抑制食品腐败,已成为食品保鲜领域的研究热点之一。
近年来,研究学者在水果、蔬菜和中草药等天然植物的提取物中筛选到很多安全、有效的活性成分,这些物质作为群体感应抑制剂可有效降低微生物的毒性[3-5]。大蒜提取物是具有群体感应抑制活性的物质,能在亚抑菌浓度下阻断N-酰基高丝氨酸内酯(AHLs)信号分子介导的群体感应系统,抑制其毒力因子产生[6-9],然而,目前关于大蒜提取物对腐败菌腐败能力的干扰作用仍少有报道。
本试验以在发酵鱼糜中筛选到的腐败菌维氏气单胞菌为对象,分析其AHLs 介导的群体感应现象,探究大蒜提取物对该细菌产AHLs 活性、生物膜形成能力、毒力基因表达和致腐能力的干扰作用,以期利用大蒜提取物等群体感应抑制剂作为食品保鲜剂,提高食品的安全性和货架期。
1 材料与方法
1.1 材料与试剂
材料:新鲜大蒜,购于杭州市潮王路菜市场;鱼糜,A 级海产铜盆鱼糜,由上虞市傣之味食品有限公司提供,贮藏于-18 ℃冰箱中,备用。
菌种:维氏气单胞菌,分离于发酵鱼糜,添加20%甘油,保存于本实验室-80 ℃冰箱中。根癌农杆菌【Agrobacterium tumefaeiens A136(pCF218/PCF372)】,壮观霉素抗性,用量为50 μg/mL。
试剂:甲酸、乙酸乙酯、二甲基亚砜(DMSO)、甲苯为分析纯级,国药集团化学试剂有限公司;5-溴-4-氯-3-吲哚-β-D-半乳糖苷(X-gal)、结晶紫、吖啶橙、壮观霉素、细菌基因组DNA 快速抽提试剂盒、细菌总RNA 快速抽提试剂盒,生工生物工程(上海)股份有限公司;RP-C18 F254s 反相薄层层析板,德国Merck 公司;AHLs 标准品,美国Sigma 公司。
1.2 仪器与设备
SpectraMax M5 酶标仪,美国Molecular Devices 公司;LifeEco PCR 仪,杭州博日科技有限公司;FV300 激光共聚焦显微镜,日本OLYMPUS 公司;StepOne 型荧光定量PCR 仪,美国ABI 公司;PHS-3C 型数显酸度计,上海精科仪器有限公司;K9840 自动凯氏定氮仪,济南海能仪器股份有限公司;e2695 型高效液相色谱仪,美国Waters 公司;YXQ-LS-SⅡ立式压力蒸汽灭菌锅,上海博迅实业有限公司医疗设备厂。
1.3 试验方法
1.3.1 细菌产AHLs 分子检测 参照Ravn 等[10]的方法,采用生物报告菌法分析细菌产AHLs 分子的活性和种类。
采用平行划线法分析AHLs 的产生情况,在含1%琼脂的LB 平板上涂布40 μL X-gal(20 mg/mL),将活化12 h 的根癌农杆菌与待测菌平行划线,于30 ℃中培养24 h,观察根癌农杆菌的颜色变化。
将待测菌培养液离心(8 000×g,5 min,4 ℃),上清液用等量酸化乙酸乙酯(0.5%甲酸)提取2~3次,合并有机相,旋转蒸发仪蒸干溶剂,用DMSO重新溶解,于-20 ℃保存待用。
采用平板扩散法分析AHLs 活性变化,100 mL 含1%琼脂的LB 培养基中加入4~10 mL 活化的根癌农杆菌和500 μL X-gal(20 mg/mL),混合均匀后倾注平板,待冷却凝固后打孔,加入40 μL待测菌AHLs 提取液,于30 ℃中培养36~48 h,观察平板的颜色变化。
采用薄层色谱法(TLC)分析细菌产生AHLs种类,取1~10 μL AHLs 标准品与细菌AHLs 提取物样品,点样于C18 反相薄层层析板。用层析液甲醇-水(体积比60/40)充分展开,取出后在常温下挥干溶剂。制备含根癌农杆菌的固体培养基,倾于板上,凝固后在30℃无菌密闭容器中培养36~48 h,观察颜色变化。AHLs 标准品的浓度分别为:0.1 mmol/L N-丁酰基-高丝氨酸内酯(C4-HSL)、10 μmol/L N-己酰基-高丝氨酸内酯(C6-HSL)、0.04 μmol/L N-庚酰基-高丝氨酸内酯(C7-HSL)、0.06 μmol/L N-辛酰基-高丝氨酸内酯(C8-HSL)、0.8 nmol/L 3-氧代-己酰基-高丝氨酸内酯(oxo-C6-HSL)和1.6 nmol/L 3-氧代-辛酰基-高丝氨酸内酯(oxo-C8-HSL)。
1.3.2 细菌的生长曲线及培养液pH 值的测定将活化12 h 的待测菌接种到100 mL LB 液体培养基中,在30 ℃下摇床培养24 h,间隔一定时间取样,参照GB/T 4789.2-2008《食品卫生微生物学检验 菌落总数测定》计数,同时利用酸度计测定培养液pH 值的变化。
1.3.3 基因分析 采用PCR 技术分析细菌的luxRI 同源基因、鸟氨酸脱羧酶基因(ODC)、赖氨酸脱羧酶基因(LDC)、鞭毛基因(fla)、弹性蛋白酶(ahyB)、丝氨酸蛋白酶基因(ser)、脂肪酶基因(lip)。将细菌过夜活化,利用细菌DNA 提取试剂盒提取细菌DNA 模板,使用的引物见表1。50 μL反应体系,PCR 扩增程序为:预变性95 ℃3 min,变性95 ℃30 s,退火55 ℃1 min,延伸72 ℃1 min(35 个循环),终延伸72 ℃10 min。其中,扩增ahyB 和fla 基因时退火温度为59 ℃。将扩增到的基因片段进行1%琼脂糖凝胶电泳,在紫外光下成像,同时送往生工生物工程有限公司测序。将得到的序列进行BLAST 搜索和同源性分析。

表1 PCR 所使用的引物序列Table 1 Primers used for PCR
1.3.4 大蒜提取物的制备 参照Rasmussen 等[6]的方法制备大蒜提取物。将市售大蒜去皮,称取150 g 于300 mL 甲苯中匀浆,萃取12 h 后用Whatman 1 号滤纸过滤。向滤液中加入150 mL 无菌水,在室温条件下搅拌24 h,两相分离得水相,经0.22 μm 滤膜过滤后得1 g/mL 大蒜提取物。通过二倍稀释法确定大蒜提取物对待测菌的最小抑菌浓度。
1.3.5 β-半乳糖酶活性的测定 将待测菌过夜活化12 h,取500 μL 细菌培养液接种至100 mL LB液体培养基中,分别加入1 mL 不同质量浓度的大蒜提取物(终质量浓度为0.15~1.20 mg/mL),以无菌生理盐水为对照,于30 ℃摇床中振荡培养12 h,将培养液离心(8 000×g,10 min)得上清液。参照Miller[11]的方法测定生物报告菌的β-半乳糖酶活性。
1.3.6 生物膜形成能力的测定 参照Kusada等[12]的方法,采用结晶紫染色法分析大蒜提取物对细菌生物膜形成能力的影响。将细菌培养液按1∶1 000 的比例稀释,加入无菌96 孔板中,加入不同浓度的大蒜提取物(终质量浓度为 0.15~1.20 mg/mL),30 ℃静置培养24 h。移除培养液,用无菌去离子水冲洗3 次,无菌风干燥,加入200 μL 0.1%结晶紫染色液,室温染色15 min,无菌水冲洗3 次,加入1 mL 95%乙醇漂洗生物膜,将洗液于波长570 nm 处测定OD 值。
参照Packiavathy 等[13]的方法,利用激光共聚焦显微镜(CLSM)分析细菌生物膜结构的变化。将细菌培养液按1%的比例接入含1 mL LB 培养基的24 孔板中,加入1.20 mg/mL 大蒜提取物和直径为1 cm 的圆形盖玻片,静置培养24 h。用无菌水轻轻冲洗玻片上未粘附的细胞,0.1%吖啶橙染色1 min。洗去多余的染色液,将盖玻片置显微镜下观察。该显微镜装有激发波长为515~560 nm 的激发滤光器及60×(60 倍)的油镜,其荧光显微镜照片与光镜照片均由Fluoview v5.0 软件获得。
1.3.7 基因表达量的测定 采用实时荧光定量PCR(RT-PCR)技术。细菌活化后加入1.20 mg/mL大蒜提取物培养24 h,利用细菌RNA 提取试剂盒提取细菌RNA。在20 μL 反应体系中,加入10 μL 2×荧光定量PCR 预混体系,0.4 μL 10 μmol/L上、下游引物,2 μL cDNA 模板。PCR 扩增程序为:预变性95 ℃3 min,变性95 ℃7 s,退火57 ℃10 s,延伸72 ℃15 s(45 个循环)。相对表达量(RQ)由2-ΔΔct方法计算得出。

表2 RT-PCR 所使用的引物序列Table 2 Primers used for RT-PCR in this study
1.3.8 细菌腐败能力的测定 参照Zhao 等[14]的方法制备无菌鱼糜肉汁。冷冻鱼糜解冻后,按质量比1∶1 加水打浆,加热煮沸5 min 后静置,冷却至室温。用2 层纱布过滤得鱼糜肉汁,加入1.6 g/L氧化三甲胺、40 mg/L L-半胱氨酸和40 mg/L L-甲硫氨酸,混合均匀后于121 ℃高压灭菌15 min,得无菌鱼糜肉汁。无菌条件下,在鱼糜肉汁中添加不同质量浓度的大蒜提取物(终质量浓度为0.15~1.20 mg/mL)。将制备好的细菌菌悬液接种至无菌鱼糜肉汁中,于30 ℃中摇床培养48 h,每间隔6 h取样,测定鱼糜肉汁中TVB-N 和腐胺含量变化。
TVB-N 的测定参照赵丹丹等[15]的方法,取5 mL 样品,加入15 mL 0.6 mol/L 高氯酸溶液均质混匀,上清液用半自动凯氏定氮仪测定。腐胺含量的测定参照Zhao 等[14]的方法,取2 mL 样品,加入25 mL 0.4 mol/L 高氯酸溶液均质混匀,上清液用丹磺酰氯衍生后用高效液相色谱分析。
1.4 数据统计分析
运用SPSS 21.0 软件和Origin 8.5 软件分析数据。测定结果以均值±标准差(n ≥3)表示,采用最小显著差异法(LSD)进行显著性差异分析,显著性水平为5%。
2 结果与分析
2.1 维氏气单胞菌产AHLs 信号分子的种类
以根癌农杆菌为报告菌,利用平行划线法分析维氏气单胞菌的AHLs 分子产生情况,结果见图1。细菌产生的AHLs 可通过琼脂扩散诱导根癌农杆菌产生β-半乳糖苷酶,并分解底物X-gal产生蓝色色素。由图1 可知,维氏气单胞菌可产生AHLs 类信号分子并诱导根癌农杆菌显蓝色。
根癌农杆菌对不同碳链长度或带有不同基团的AHLs 分子敏感程度不同,这些AHLs 经TLC 展开,在培养基相应的位置诱导根癌农杆菌呈现蓝色斑点。根据斑点的比移值、形状和颜色深、浅可直观判断AHLs 的种类及相对含量。对维氏气单胞菌的AHLs 提取液的TLC 分析结果见图2。分析可知,该细菌至少产生3 种AHLs 分子,即:C6-HSL、C7-HSL 和C8-HSL。C4-HSL 与C6-HSL 是目前气单胞菌属报道最多的2 种AHLs 信号分子[16]。Li 等[17]在腐败比目鱼中分离的温和气单胞菌中检测到C4-HSL、C6-HSL、C8-HSL、N-癸酰基-高丝氨酸内酯(C10-HSL)和N-十二酰基-高丝氨酸内酯(C12-HSL)。本试验在维氏气单胞菌中检测到侧链上的碳原子数目为奇数的AHL 信号分子,即C7-HSL。
2.2 维氏气单胞菌生长过程中AHLs 活性变化
细菌产生的AHLs 分子浓度受细菌密度变化影响,不同生长阶段的细菌AHLs 提取液能诱导根癌农杆菌产生不同直径的蓝色圈。维氏气单胞菌生长过程中的细菌总数与培养液pH 值的变化见图3,AHLs 活性变化见图4。分析可知,细菌培养4 h 时处于对数生长期初期,此时细菌分泌的AHLs 可诱导根癌农杆菌显蓝色。在4~10 h 细菌的快速生长阶段,根癌农杆菌的变色直径不断增大,说明AHLs 的含量快速累积。在10~18 h 时,根癌农杆菌蓝色圈的直径最大,此时培养液中AHLs 浓度最高。在18~48 h,细菌处于生长稳定期,诱导报告菌变色的直径减小,说明AHLs 的浓度开始降低。研究发现,AHLs 含量的减少与环境pH 值升高有关[18]。AHLs 的高丝氨酸内酯环在碱性条件下结构不稳定,易解环[19]。对细菌生长过程中培养液的pH 值变化的分析表明:细菌培养18 h 后培养液的pH 值由6.92 增至8.11,培养48 h时pH 值达8.97。随着细菌的生长,培养液的pH值不断升高,AHLs 开始降解,活性降低。
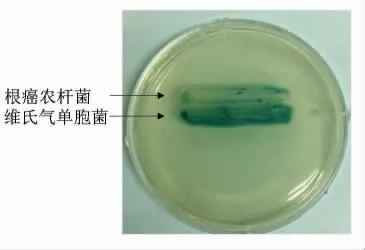
图1 利用根癌农杆菌分析维氏气单胞菌的AHLs 产生情况Fig.1 Detection of AHLs production of A.veronii bv.veronii using A.tumefaeiens A136
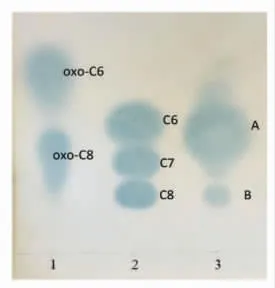
图2 维氏气单胞菌产AHLs 的种类Fig.2 The types of AHLs produced in A.veronii bv.veronii
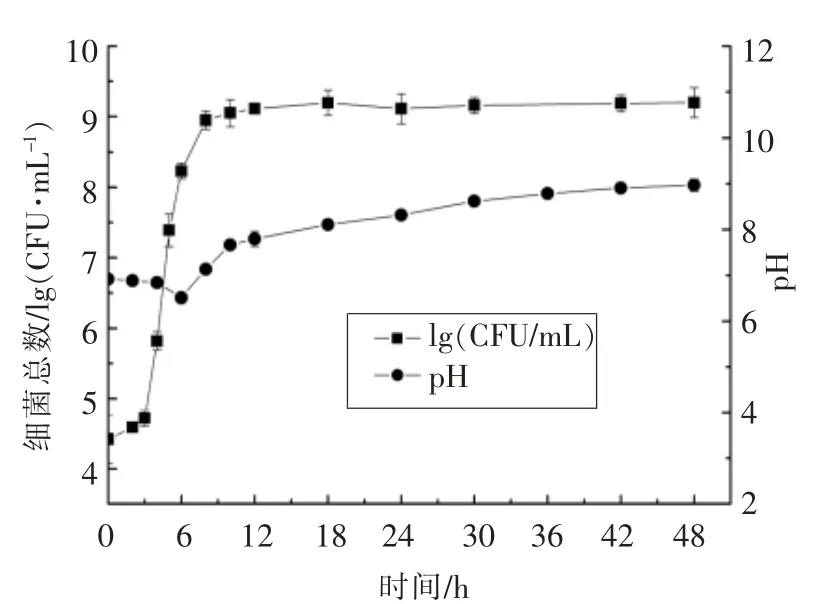
图3 维氏气单胞菌生长过程中的细菌总数与pH 值的变化Fig.3 The changes in growth number and pH values of A.veronii bv.veronii during growth
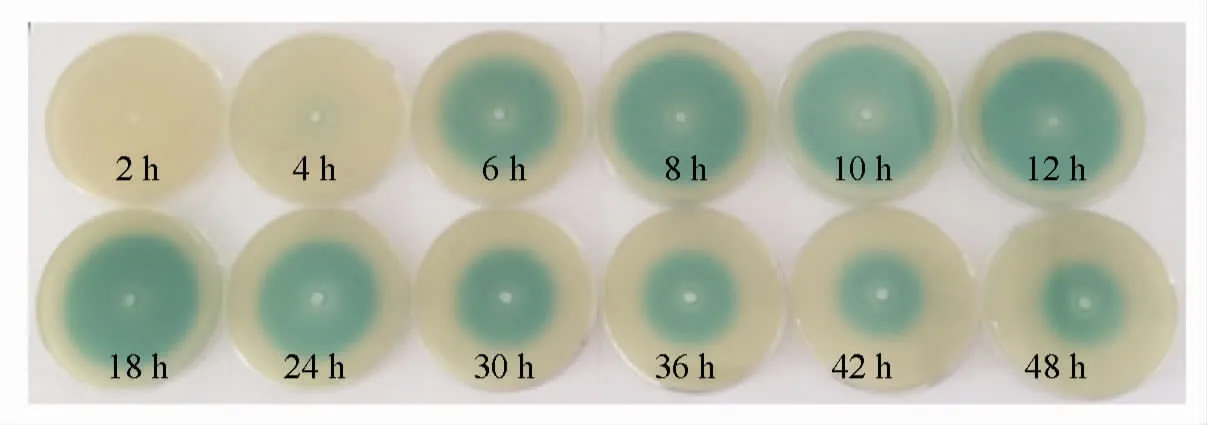
图4 维氏气单胞菌在生长过程中AHLs 的活性变化Fig.4 The changes in AHLs activities of A.veronii bv.veronii during growth
2.3 维氏气单胞菌的基因分析
革兰氏阴性菌AHLs 介导的群体感应系统主要由luxRI 的同源基因调控[20]。利用PCR 技术分析维氏气单胞菌中的luxRI 同源基因,结果见图5a。分析发现2 个片段序列分别为转录调控因子acuR 和自诱导基因acuI,这与Jangid 等[21]的研究结果相同,该学者首次在维氏气单胞菌MTCC 3249 中检测到acuRI 群体感应系统。此外,分析维氏气单胞菌的氨基酸脱羧酶基因和毒力基因,结果见图5b、5c。鸟氨酸脱羧酶基因(ODC)和赖氨酸脱羧酶基因(LDC)的检出说明该细菌具有产腐胺和尸胺的能力,胞外酶基因(ahyB,ser,lip)的检出说明该菌具有一定的蛋白酶活性和脂肪酶活性,鞭毛基因(fla)的检出说明该菌具有鞭毛运动和形成生物膜的能力。
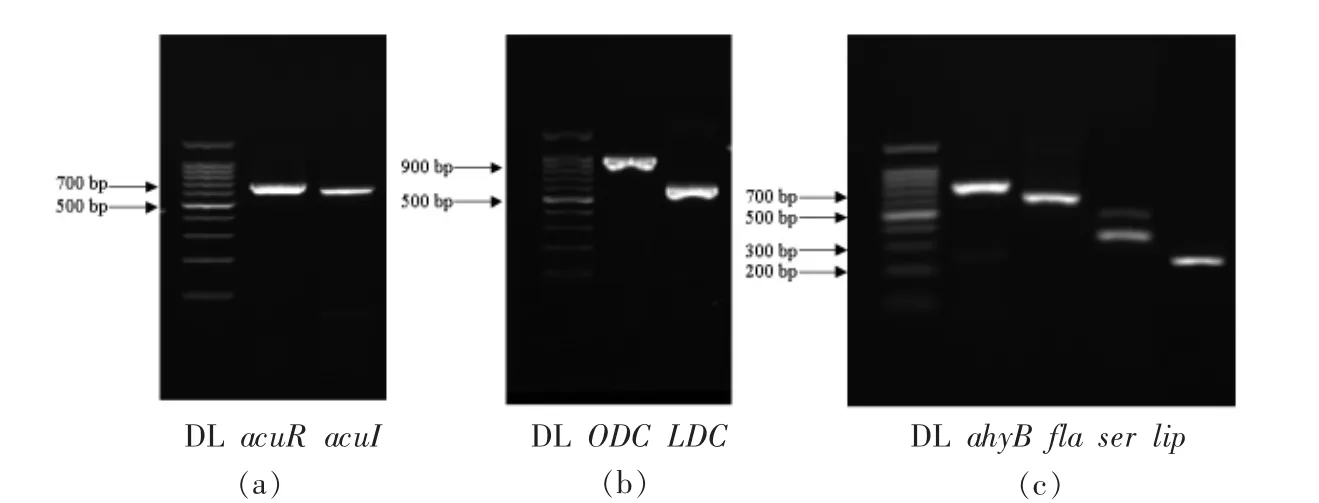
图5 维氏气单胞菌基因的PCR 产物电泳图Fig.5 Agarose gel electrophoresis of PCR products of genes in A.veronii bv.veronii
2.4 大蒜提取物对维氏气单胞菌产AHLs 的抑制作用
大蒜提取物对维氏气单胞菌的最小抑菌质量浓度为2.5 mg/mL(数据未显示)。本试验选择低于此质量浓度的系列(0,0.15,0.3,0.6,1.2 mg/mL)来研究大蒜提取物对维氏气单胞菌群体感应的干扰作用。
通过检测生物报告菌根癌农杆菌的β-半乳糖苷酶活性来分析不同浓度大蒜提取物对维氏气单胞菌产生AHLs 活性的干扰作用,结果见图6。分析可知,随着大蒜提取物质量浓度的增加(0~1.20 mg/mL),根癌农杆菌的β-半乳糖苷酶活性不断降低(P<0.05)。这说明大蒜提取物对维氏气单胞菌产AHLs 活性具有显著的抑制作用(P<0.05),且大蒜提取物的质量浓度与其群体感应抑制活性呈明显量效关系。大蒜提取物质量浓度为1.20 mg/mL 时对根癌农杆菌的β-半乳糖苷酶活性的抑制率最高,为39.6%。这可能是因为此质量浓度下维氏气单胞菌AcuR 蛋白与群体感应抑制活性物质的结合力最强,抑制了AHLs 分子的转录表达[22]。
2.5 大蒜提取物对维氏气单胞菌生物膜形成能力的干扰作用
气单胞菌属的成膜能力是受群体感应调控的。通过结晶紫染色法和CLSM 技术分析大蒜提取物对维氏气单胞菌生物膜形成能力的影响,结果见图7、图8。分析可知,大蒜提取物对维氏气单胞菌的生物膜形成有显著的抑制作用(P<0.05),且大蒜提取物的质量浓度与其抑制活性呈量效关系。与对照组相比,在1.20 mg/mL 大蒜提取物的作用下,细菌培养24 h 后形成的生物膜量显著降低36.7%(P<0.05)。通过分析维氏气单胞菌成膜结构的变化,发现经1.20 mg/mL 大蒜提取物处理,该细菌形成的生物膜细胞密度明显减小,结构较为疏松。Li 等[9]发现大蒜精油中的群体感应活性物质能有效抑制铜绿假单胞菌的生物膜产生及膜中的细菌数量。Bjarnsholt 等[7]研究发现,大蒜提取物处理后铜绿假单胞菌形成的生物膜对抗生素的耐受性大大降低。利用群体感应抑制剂干扰食源性腐败菌或病原菌的生物膜形成,可降低微生物污染,提高食品加工的安全性。
2.6 大蒜提取物对维氏气单胞菌基因表达量的影响
利用RT-PCR 技术分析大蒜提取物对维氏气单胞菌基因表达量的影响,结果见图9。与对照组相比,经1.20 mg/mL 的蒜提取物处理,acuR、acuI、ODC、fla、ahyB、ser 的基因表达量均显著下调(P<0.05),LDC 和lip 的基因表达量没有显著性变化(P>0.05)。其中,acuR 和acuI 基因的表达量分别减少32.8%和34.6%,说明维氏气单胞菌的AHLs群体感应表达受到抑制。ODC 基因表达量的下调说明该菌产腐胺的能力受到一定程度的抑制。ahyR 和ser 基因表达量的下调,说明维氏气单胞菌胞外蛋白酶活性受到抑制。Rasmussen 等[6]研究发现,经20 mg/mL 大蒜提取物处理,铜绿假单胞菌的弹性蛋白酶基因的表达量下调10 倍。此外,维氏气单胞菌经1.20 mg/mL 大蒜提取物处理,鞭毛基因fla 的表达量下调37.2%。Myszka 等[23]研究发现,通过干扰细菌群体感应并抑制鞭毛基因表达,能显著降低细菌在不锈钢表面的聚集程度和成膜能力。
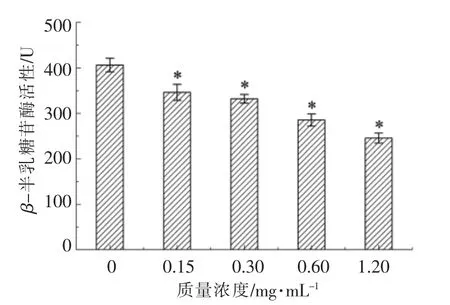
图6 不同质量浓度大蒜提取物对β-半乳糖苷酶活性的影响Fig.6 The effects of garlic extracts of different mass concentrations on β-galactosidase assay
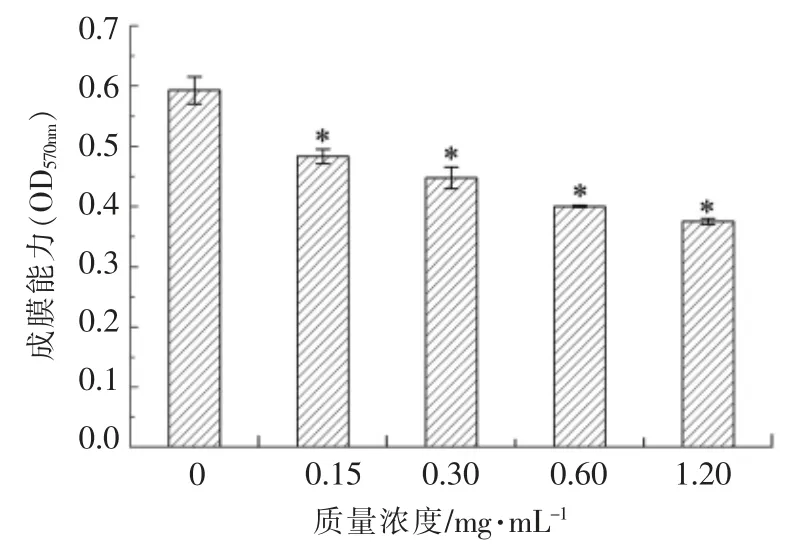
图7 不同质量浓度大蒜提取物对维氏气单胞菌成膜能力的影响Fig.7 The effects of garlic extract at different mass concentrations on biofilm formation in A.veronii bv.veronii
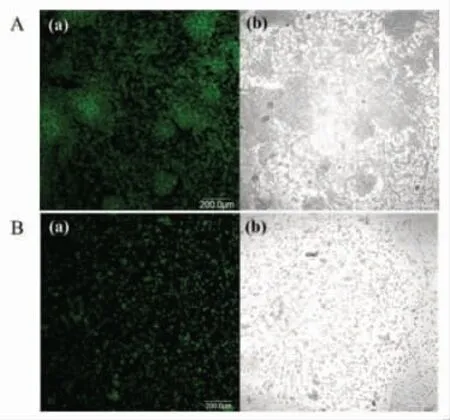
图8 1.20 mg/mL 大蒜提取物对维氏气单胞菌生物膜结构影响的CLSM 图Fig.8 The CLSM images of biofilm structure of A.veronii bv.veronii grown in the absence of 1.20 mg/mL garlic extract
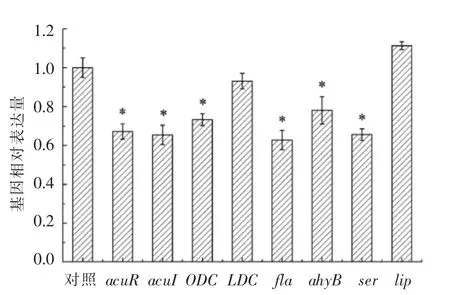
图9 1.20 mg/mL 大蒜提取物对维氏气单胞菌基因表达量的影响Fig.9 The effects of 1.20 mg/mL garlic extract on gene expression in A.veronii bv.veronii
2.7 大蒜提取物对维氏气单胞菌腐败能力的干扰作用
腐败菌产TVB-N 的能力与其分泌的胞外蛋白酶活力有关。细菌的蛋白酶活力越高,产TVBN 能力越强。大蒜提取物对接种维氏气单胞菌的鱼糜肉汁TVB-N 含量的影响见图10。可见,大蒜提取物对维氏气单胞菌产生TVB-N 的能力具有一定抑制作用,且呈明显的量效关系,其中1.20 mg/mL 大蒜提取物对细菌产TVB-N 的抑制作用最为显著(P<0.05)。培养36 h 后,含1.20 mg/mL大蒜提取物的鱼糜肉汁的TVB-N 含量为24.06 mg/100 mL,而对照组中TVB-N 含量为28.56 mg/100 mL,即经1.20 mg/mL 大蒜提取物处理,鱼糜肉汁中TVB-N 的含量降低15.8%。这可能与维氏气单胞菌的胞外蛋白酶基因表达受到抑制有关。
维氏气单胞菌产腐胺的能力与其分泌的鸟氨酸蛋白酶活力有关,酶活力越高,细菌产腐胺的能力越强。大蒜提取物对接种维氏气单胞菌的鱼糜肉汁腐胺含量的影响见图11。可见,大蒜提取物对维氏气单胞菌在鱼糜肉汁中产腐胺的能力具有一定抑制作用。培养36 h 后,对照组的腐胺含量为144.03 μg/mL,1.20 mg/mL 大蒜提取物处理组的鱼糜肉汁腐胺含量为118.51 μg/mL,降低17.7%。这可能是因为大蒜提取物干扰维氏气单胞菌的鸟氨酸脱羧酶基因的表达,抑制其鸟氨酸蛋白酶活性。
群体感应抑制剂可有效降低食源性腐败菌的毒性和致腐能力,通过该抑制剂干扰腐败菌的群体感应表达是防止食品腐败的有效途径。Vittal[24]研究了咖喱树叶精油对腐败菌假单胞菌群体感应系统的干扰作用,发现在亚抑菌浓度范围,该精油能够延缓冷藏牛奶的腐败现象。Li 等[25]发现0.1 μL/mL 肉桂醛可干扰腐败菌的群体感应表达并延长比目鱼片的冷藏货架期。Gui 等[26]发现6.6 U/mL AHL 内酯酶能降低维氏气单胞菌的蛋白酶活力,延长冷冻鲟鱼的货架期。本研究中,大蒜提取物使维氏气单胞菌的胞外蛋白酶及氨基酸脱羧酶的基因表达下调,减少鱼糜肉汁中TVB-N 和腐胺含量的累积,说明大蒜提取物通过干扰维氏气单胞菌的群体感应表达降低其腐败能力,延缓鱼糜制品的腐败。
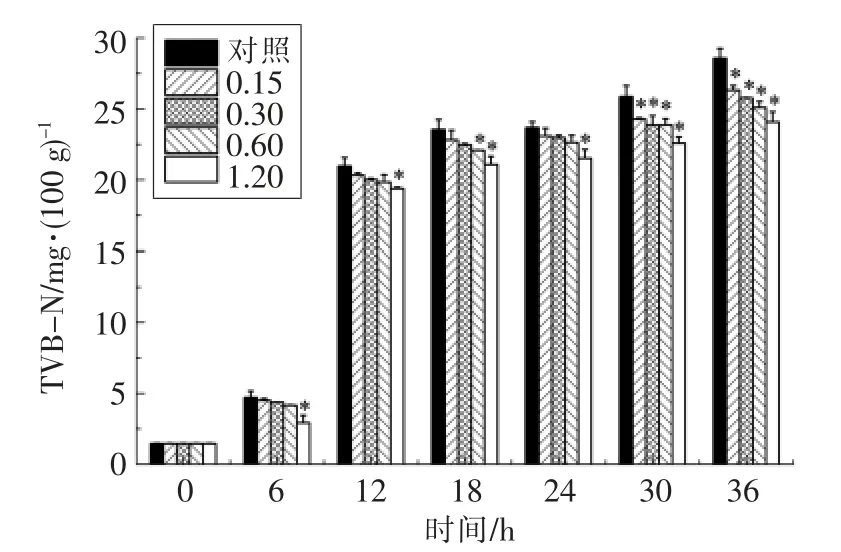
图10 1.20 mg/mL 大蒜提取物对维氏气单胞菌产TVB-N 能力的影响Fig.10 The effects of 1.20 mg/mL garlic extract on the production of TVB-N in A.veronii bv.veronii
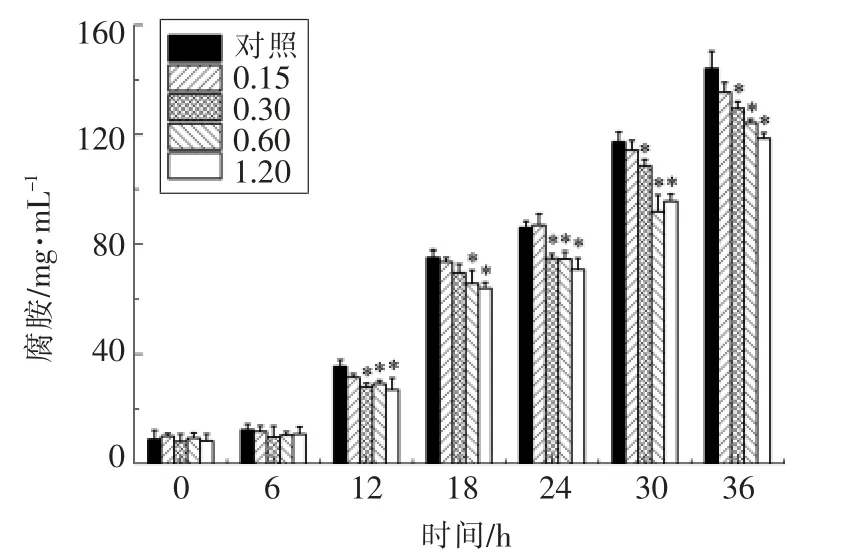
图11 1.20 mg/mL 大蒜提取物对维氏气单胞菌产腐胺能力的影响Fig.11 The effects of 1.20 mg/mL garlic extract on the production of putrescine in A.veronii bv.eronii
3 结论
本研究发现,发酵鱼糜中分离的维氏气单胞菌具有AHLs 介导的群体感应现象,可产生C6-HSL、C7-HSL 和C8-HSL,且AHLs 分子活性受细菌生长密度和培养液pH 值的影响呈先增加后降低的趋势。大蒜提取物对维氏气单胞菌的AHLs活性、成膜能力、群体感应调控基因表达及腐败能力的影响是:通过干扰细菌AHLs 介导的群体感应表达,降低细菌的毒性和腐败能力。在今后的研究中,可对大蒜提取物进行分离纯化,研究其活性成分对鱼糜制品腐败变质的调控作用。
