基于GC-MS分析东北红豆杉野生种与栽培种的代谢差异
2021-03-02刘龙桀吴可心刁云飞李中跃穆立蔷
刘龙桀 吴可心 刁云飞 李中跃 穆立蔷*
(1. 东北林业大学林学院,哈尔滨 150040;2. 黑龙江牡丹江森林生态系统定位研究站,哈尔滨 150081)
东北红豆杉(Taxus cuspidataSieb.et Zucc.)又名紫杉,是红豆杉科(Taxaceae)红豆杉属(Taxus)植物[1]。主要分布我国黑龙江和吉林等地区,朝鲜、日本也有少量分布。东北红豆杉是第三纪冰川时期孑遗植物[2],是东北地区唯一的极小种群,东北红豆杉存在生境狭小[3]、分布地不连接等问题,属于国家Ⅰ级珍稀保护植物[4]。东北红豆杉幼苗在第一年生长较慢,之后生长增速,每年可达20~30 cm[5]。因为野生幼苗容易遭受外界破坏导致死亡率高[6],进而东北红豆杉野生种群更新率低。从东北红豆杉树皮、枝、叶中所提取的紫杉醇[7]是目前世界上公认对治疗乳腺癌、皮肤癌、卵巢癌和肺癌具有特殊的疗效的药品[8]。随着对东北红豆杉的进一步研究,在东北红豆杉提取液中提取出次级代谢产物倍半萜类、木质素类、黄酮类、生物碱化合物和挥发性油脂类更具有医疗前景[9]。次级代谢产物黄酮类化合物具有多种药理作用,如抗氧化、抗病毒、降血压、降血脂、抗炎、保护心脏和抗疲劳等作用[10~13]。还可以当作保健品的主要成分具备良好的保健功效[13]。近年来国家对东北红豆杉的保护力度加大,市场对紫杉醇抗癌类药物的需求量也在增大,在进一步的研究中发现东北红豆杉的嫩枝中也存在紫杉醇、三尖杉宁碱、紫杉碱和黄酮类化合物[14]。虽然人工栽培东北红豆杉也逐渐规模化、专业化[15],但对其的研究还比较少,不具体。
代谢组学主要是将动植物体内的代谢产物进行定量、定性分析,是通过高通量检测和统计学数据分析的方法来进行对动植物体内代谢规律探索的一门学科。通常的检测手段有核磁共振、GCMS、LC-MS 和稳定性同位素等方法。代谢组学在研究红豆杉方向应用也比较广泛,像在代谢层次上来分析红豆杉中紫杉醇的合成途径[16],代谢组学还可以鉴定紫杉烷类化合物[17]。陈一鸣[18]通过利用LC-MS 研究了南方红豆杉(Taxus wallichianavar.mairei)野生种和观赏栽培种金锡杉代谢差异,发现野生种和栽培种在糖类和黄酮类化合物代谢上差异显著。李影影[19]通过测定光照对南方红豆杉对初生代谢产物,发现光照对南方红豆杉初生代谢产物有明显的影响。张春椿[20]通过利用GCMS 对南方红豆杉种皮抑制物进行鉴定分析,主要成分有十六烷酸、十八烷酸、9-十八碳烯酸、9,12-十八碳二烯酸等。初生代谢产物不仅对植物生理活动有重要的影响,对植物组织和细胞也有明显的作用。东北红豆杉作为抗癌药物的原材料,国内主要对次级代谢物质研究较为深入,但通过GCMS 技术对东北红豆杉野生种与栽培种的初级代谢产物进行分析的报道较少,本实验通过非靶向代谢组学来探究东北红豆杉野生种与栽培种的树皮,来获取差异代谢物和相关代谢通路的信息,来揭示野生东北红豆杉与栽培种的代谢差异,为今后东北红豆杉进一步开发和东北红豆杉野生种的保护提供理论依据。
1 材料与方法
1.1 材料与试剂
植物样本于2019 年9 月采自黑龙江省穆棱市东北红豆杉国家级自然保护区和东北林业大学树木园,选择相同径级生长情况相同的雌性东北红豆杉各3 棵,每个植物分别采集3 个生物学重复,经东北林业大学穆立蔷教授鉴定均为东北红豆杉,将其洗净后自然晾干,备用。甲醇(色谱级,CNW Technologies),吡啶(分析纯,Adamas),甲氧铵盐(TCI),核糖醇(纯度≥99%,SIGMA),BSTFA(with 1% TMCS,v/v),衍生试剂(REGIS Technologies),饱和脂肪酸甲酯(Dr.Ehrenstorfer)纯水。
1.2 仪器与设备
7890A 气相色谱(安捷伦公司,美国),PEGASUSHT 质谱仪(LECO,美国),Heraeus Fresco17 高速离心机(赛默飞世尔科技公司,美国),Forma 900 series 超低温冰箱(赛默飞世尔科技公司,美国),BSA124S-CW 分析天平(赛多利斯公司,德国),JXFSTPRP-24 研磨仪(上海净信科技有限公司),YM-080S 超声仪(深圳市方奥微电子有限公司),DHG-9023A 烘箱(上海一恒科学仪器有限公司),LNG-T98 真空干燥仪(太仓市华美生化仪器厂)。
1.3 样品的制备
称取东北红豆杉野生种和栽培种样本50±1 mg 于2 mL 离心管中,加入500 μL 预冷提取液(甲醇水体积比=3∶1),再加入10 μL 核糖醇,涡旋30 s;加入瓷珠,放入45 Hz 研磨仪处理4 min,取出后继续超声5 min(冰水浴);将样本4℃下12 000 r·min-1离心15 min;小心移取30 μL 上清液于1.5 mL EP 管中,每个样本各取20 μL 混合成QC样本;将样本放在真空浓缩器中干燥提取物;向干燥后的代谢物加入100 μL 甲氧胺盐试剂(甲氧胺盐酸盐,溶于吡啶20 mg·mL-1),轻轻混匀后,放入烘箱中80℃孵育30 min;向每个样品中加入100 μL BSTFA(含有1% TMCS,v/v),将混合物70℃孵育1.5 h;冷却至室温,向混合的样本中加入5 μL FAMEs(溶于氯仿);进行GC-MS代谢组学分析。
1.4 GC-MS分析条件
使用安捷伦7890气相色谱仪和飞行时间质谱仪进行GC-MS 分析。系统采用DB-5MS 毛细管柱,以无分流模式注入1 μL 小份样品。使用氦气作为载气,前部入口吹扫流量为3 mL·min-1,通过色谱柱的气体流速为1 mL·min-1。初始温度50℃保 持1 min,然 后 以10℃·min-1的 速 率 升 高 至310℃,然后在310℃保持8 min。注入、传输线和离子源温度分别为280、280和250℃。在电子碰撞模式下,能量为-70 ev。在溶剂延迟6.35 min 后,以每秒12.5 个光谱的速率,在m/z 范围为50~500 的全扫描模式下获得质谱数据。
1.5 数据处理
将原始数据使用ChromaTOF 软件(V4.3x,LECO)对质谱数据进行了峰提取、基线矫正、解卷积、峰积分、峰对齐等分析[21]。对物质定性工作中,使用了LECO-FiehnRtx5数据库,包括质谱匹配及保留时间指数匹配。最后,将QC 样本中检出率50%以下或RSD>30%的峰去除[22]。导出数据至Excel 进行后续整理,使用SIMCA 软件(V16.0.2,Sartorius Stedim Data Analytics AB,Umea,Sweden),对数据进行对数(LOG)转换加中心化(CTR)格式化处理,然后进行自动建模分析。
2 结果与分析
2.1 主成分分析
为了将东北红豆杉野生种和栽培种在代谢物积累上的差异科学的辨别出来,首先使用统计学上的主成分分析法进行分析,PCA 可以揭示数据的内部结构,从而更好的解释数据变量[23]。使用SIMCA 软件(V16.0.2,Sartorius Stedim Data Analytics AB,Umea,Sweden)进行分析[24],得到了一组拟合了两个主成分的PCA 模型,累计R2X=0.741,模型参数数值均大于0.5,且二者的数值都在置信区,在合理可以接受范围内,说明根据实验建立PCA 模型稳定,可以用于代谢组学实验说明以及进行代谢差异分析。从图1 中可以得到东北红豆杉野生种都在坐标轴的负方向,而栽培种在坐标轴的正方向上,二者分别位于坐标轴的两侧被分开,说明东北红豆杉野生种和栽培种之间的代谢物存在明显的不同。但PCA 分析法存在缺点,首先主成分分析中的主成分是模糊的,并不能很好的解释主成分内容之间的关系,其次小成分也可能起到特殊作用,用于区分样本之间的差异性,而且主成分分析法没有监督性。
2.2 OPLS-DA模型的分析结果
2.2.1 OPLS-DA
OPLS 又叫正交偏最小二乘法判别分析,它是多因变量对多自变量的回归建模方法,其特点是可以除去自变量X 中与分类变量Y 无关的数据因素,并可以对正交变量和非正交变量进行分别解读和分析,可以更加准确的分析代谢产物在不同样本的差异和关联。OPLS-DA 分析以变量的重要性为主要依据,对代谢化合物进行选择和甄别,更能把握多位数据整体特征和变异规律[25],使用SIMCA 软件(V16.0.2,Sartorius Stedim Data Analytics AB,Umea,Sweden)对数据进行对数(LOG)转换加UV 格式化处理,首先对第一主成分进行OPLS-DA 建模分析,然后用交叉验证后得到的R2Y 和Q2对模型有效性进行评判[26],如图2所示东北红豆杉野生种在置信区左侧,而栽培种在置信区的右侧,说明该OPLS-DA 模型能够有效的区分两种东北红豆杉植物组织样品。建立的OPLS-DA模型拥有两个主成分,RX2=0.65,RY2=0.999,Q2=0.925,这三个模型参数指数均大于0.5,符合我们对实验数据模型的预期,说明本研究所建立的OPLS-DA 模型可以有效的说明东北红豆杉野生种和栽培种的代谢产物的差异和区别。
2.2.2 OPLS-DA置换检验
OPLS-DA 模型分析是有监督分类的模型,可能会出现拟合现象。模型可以将样本很好的进行区分,但用来预测新的样本时,效果却很差,无法达到我们的预期结果。可以用置换验证这种模型评判方式来验证模型的可靠性。通过将每个样本的分组标记随机打乱,多次建立与之相对应的OPLS-DA 模型,以获取多次建立模型的Q2,一个可靠模型的Q2应该远大于随机建立模型的Q2。对OPLS-DA模型的置换验证如图3所示,右侧原始OPLSDA模型的Q2远远大于左侧随机模型的Q2,说明该OPLS-DA模型不存在过拟合现象,可靠性良好。
2.2.3 两种东北红豆杉样品差异代谢物的确定
以OPLS-DA 模型中VIP 值(VIP≥1),同时与t检验的结果P值(P≤0.05)综合分析来确定两份东北红豆杉样品的标志性差异代谢产物。由表1 可知,两份东北红豆杉样品共筛选出共45 种差异代谢产物。这些差异代谢物主要是4 种氨基酸包括鸟氨酸,瓜氨酸,天冬氨酸和O-乙酰丝氨酸,10 种有机酸包括棕榈酸,硬脂酸,4-羟基苯甲酸,阿魏酸,花生油酸,3-苯基乳酸等和一些多酚,糖醇等。其中有23种代谢物log2FC>0表示为下调,包括棕榈酸,4-羟基苯甲醛,花生油酸,二氢香豆素,瓜氨酸,鸟氨酸,2,4-二氨基丁酸,环己基甲酰胺。其他22种差异代谢物质log2FC<0表示为上调,包括龙胆二糖,D-阿拉伯糖醇,来苏糖,表儿茶素,核糖等。大多数的氨基酸和有机酸表示为下调,氨基酸和有机酸均为主要抗逆调节物质,缓解植物在逆境的影响。说明不同生长方式下的东北红豆杉,初级代谢产物积累存在差异。
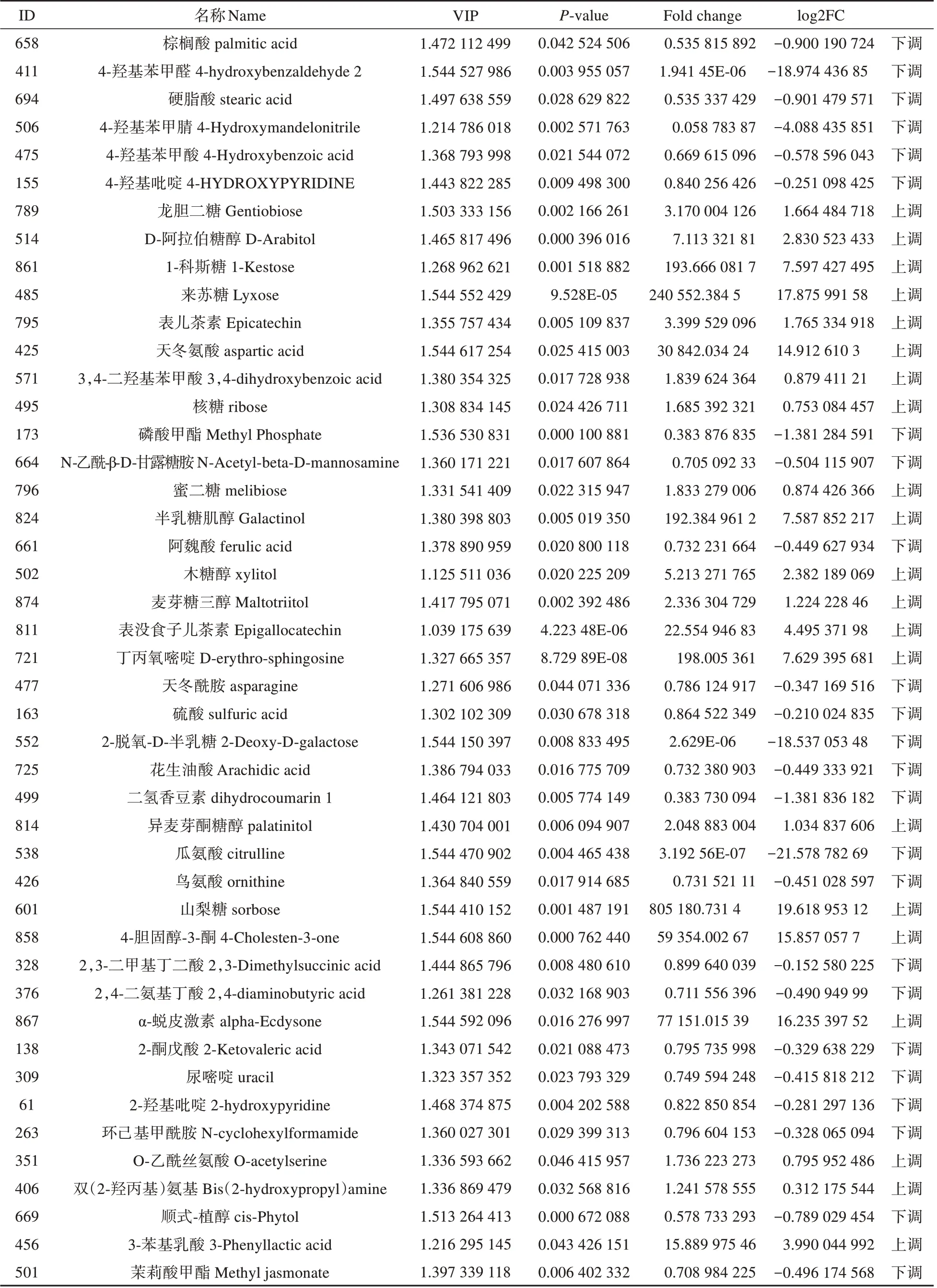
表1 筛选的差异代谢物Table 1 Screening of differential metabolites
2.3 差异代谢物分析
2.3.1 差异代谢物通路分析
通过查找KEGG 数据库可以得到差异代谢化合物的ID,将获得到的差异代谢物ID 输入到KEGG 代谢通路数据库,进行代谢富集分析[27],将两者可能存在的差异代谢途径甄选出来,对这些代谢途径和差异代谢物再进行相关性分析[28],总共检测到22 条代谢通路。将结果绘制成气泡图,见图4,气泡大小与impact 值有关,气泡越大impact 值越大,气泡越小,impact 值越小。红色的颜色越深代表-ln(p)的值越大,蓝色越深表示-ln(p)的值越小,最后通过impact 值和-ln(p)值综合分析得到与代谢物差异最相关的代谢途径。如图4 所示,代谢物差异的代谢途径共有7 条比较显著,分别为硫代谢,精氨酸和脯氨酸代谢,磷酸戊糖途径,不饱和脂肪酸的生物合成,半乳糖代谢,丙氨酸、天冬氨酸和谷氨酸代谢以及苯丙烷类生物合成。其中影响最明显的是硫代谢,在该代谢通路检测到的差异代谢物O-乙酰丝氨酸,该通路与蛋白质的合成密切相关。其次是精氨酸和脯氨酸代谢与丙氨酸、天冬氨酸和谷氨酸代谢,两条都是氨基酸代谢,参与的氨基酸与有机酸可以减轻逆境对植物的有害影响,对维护植物相对稳定的状态起到一定作用,说明东北红豆杉通过氨基酸和有机酸的增加和代谢通路的调节来降低环境胁迫对植物细胞和组织的影响,提高对环境的适应性和抵抗性。东北红豆杉野生种除天冬氨酸表达上调外,代谢通路所涉及的其他三种氨基酸均表达下调,所以东北红豆杉的野生种和栽培种相比,栽培种的氨基酸和有机酸呈上调,栽培种的适应性和抵抗性更强。
2.3.2 差异化合物通路分析
7 种差异代谢途径大体上分成三种。第一种类型氨基酸和有机酸代谢途径,包括硫代谢,精氨酸和脯氨酸代谢,不饱和脂肪酸的生物合成,丙氨酸、天冬氨酸和谷氨酸代谢。这些代谢途径都是与蛋白质,有机酸合成有关。脯氨酸,鸟氨酸可以在干旱逆境对植物细胞起到渗透调节作用[29],对植物抵抗外界环境变化起到一定作用。东北红豆杉栽培种比野生种更加依赖,通过合成蛋白质和氨基酸来缓解逆境所带来的影响和伤害。第二种类型磷酸戊糖途径和半乳糖代谢均是糖代谢。这两种代谢途径中的差异代谢物呈上调,说明在能量代谢方向,野生的东北红豆杉是优于栽培种的。第三种代谢途径是苯丙烷类生物合成,苯丙烷类化合物影响植物体内细胞的分化、着色色素的形成及木质素的合成,与黄酮类化合物合成有关。
3 讨论
本研究通过使用GC-MS技术和非靶向代谢组学的研究方法,研究了东北红豆杉野生种和栽培种在初级代谢方面的差异。从样品中检测出45种差异代谢物,代谢物的种类有氨基酸类、糖类、有机酸类化合物和脂肪酸类化合物等。其中氨基酸有4 种包括鸟氨酸、天冬氨酸、瓜氨酸和O-乙酰丝氨酸,糖类有半乳糖、木糖醇等,有机酸有10 种包括棕榈酸、硬脂酸、阿魏酸等。有23种差异代谢物表示为下调,其余22 种代谢物表示为上调。将差异代谢物通过代谢通路富集分析,得到22 条代谢通路。其中有7 条通路比较显著,分别为硫代谢、精氨酸和脯氨酸代谢、磷酸戊糖途径、不饱和脂肪酸的生物合成、半乳糖代谢、丙氨酸、天冬氨酸和谷氨酸代谢以及苯丙烷类生物合成。
在精氨酸和脯氨酸代谢途径中,有3种差异性代谢物参与其中,分别是鸟氨酸、瓜氨酸和天冬氨酸。除天冬氨酸外,其他两种代谢物的积累,野生东北红豆杉低于栽培种。根据刘建兵[30]研究发现,干旱胁迫下马尾松(Pinus massonianaLamb.)苗木的脯氨酸及游离氨基酸显著增加,使得苗木能在干旱环境下生长。鸟氨酸是脯氨酸的前体物质[31],脯氨酸可以对植物细胞起到渗透调节作用,在一些逆境胁迫条件下如干旱胁迫,脯氨酸可以增强植物细胞的保水和吸水能力来解决因水分匮乏而引起的渗透问题,脯氨酸对植物在重金属胁迫下起到一定的缓解作用[32],脯氨酸还有一定的抗氧化性,可以缓解重金属胁迫导致的氧化胁迫。鸟氨酸的显著积累可以促进脯氨酸的合成与积累,东北红豆杉栽培种鸟氨酸相对含量高于野生种,东北红豆杉野生种在干旱和重金属胁迫下比栽培种的抗逆性弱一些。糖代谢也是植物代谢的重要途径之一,糖代谢与植物在一些环境下的抗逆性应答以及植物体生长有关。包雨卓[33]采用GC-MS 技术分析冬小麦(Triticum aestivumL.)在低温胁迫下的代谢物质变化,发现大部分糖类、氨基酸和有机酸显著增加,且随着温度降低而升高,说明冬小麦通过大部分糖类、氨基酸和有机酸的显著增加来抵御低温胁迫,通过富集得到的差异糖代谢途径是半乳糖代谢和戊糖磷酸途径,其中戊糖磷酸途径[34]是植物体内重要的葡萄糖代谢途径之一。检测到戊糖磷酸途径中的核糖和6-磷酸葡萄糖-D-内酯的积累,东北红豆杉野生种高于栽培种。在半乳糖代谢的差异代谢产物蜜二糖积累也是东北红豆杉野生种高于栽培种,说明野生东北红豆杉在能量供应上也是强于栽培种。这可能和野生东北红豆杉所处的保护区地理环境气温急剧变化有关。穆棱保护区年平均气温-2℃极端最低气温-44.1℃,最高气温35.7℃,昼伏温差大[35]。野生东北红豆杉需要大量的能量来抵御低温胁迫。检测到的苯丙烷类代谢途径中差异代谢化合物阿魏酸,阿魏酸是苯丙烷类代谢的中间产物,与多糖,木质素构成植物细胞壁的一部分。阿魏酸具有强抗氧化性和抗菌性,可以清除植物体内的自由基和过氧化物[36]。根据方紫雯[37]的研究,阿魏酸对凤丹(Paeonia ostiiT.)干旱胁迫有一定的缓解效应,东北红豆杉栽培种比野生种有更强的抗氧化性和抗菌能力,对干旱胁迫形成氧自由基有良好的清除效果。
4 结论
本实验基于GC-MS的代谢组学技术对东北红豆杉野生种和栽培种样品进行代谢组学分析。实验共鉴定出45 种差异代谢物,发现东北红豆杉栽培种中大部分氨基酸和有机酸的积累比例高于野生种,东北红豆杉栽培种拥有更多抗氧化性物质和缓解逆境胁迫的调节物质,可以更快的缓解和消除如干旱,重金属等逆境胁迫所带来的伤害和影响,更好地保护植物组织和细胞,东北红豆杉栽培种对胁迫逆境下具有更强抗性和适应性。东北红豆杉野生种的糖类积累高于栽培种,在应对低温胁迫时有更好的抗性,并且能够更及时的缓解胁迫环境带来的有害影响,这可能和东北红豆杉野生种所处的特殊地理环境穆棱保护区年平均气温较低有关。但鉴于GC-MS的代谢组学技术仍有一定的局限性,实验对一些物质的检测存在不足,如萜类化合物没有检测到。对接下来的研究,应该运用其他检测方法如LC-MS,进行更全面,多角度的实验与探究。东北红豆杉野生种亟需更多的保护,本研究为进一步研究东北红豆杉的抗逆性及保护野生东北红豆杉奠定了理论基础。
