绿色环保肥料增效剂聚天冬氨酸对杨树幼苗生长的影响
2021-03-02王世震
曹 丹 王世震 韩 梅
(南京林业大学生物与环境学院,南方现代林业协同创新中心,南京 210037)
氮(Nitrogen,N)是植物生长和发育最重要的元素之一[1]。给杨树施加一定浓度的氮素会促使生物量、光合速率和株高等方面的显著提高。氮素的多少也直接影响杨树的材积量与胸径[2~t4]。杨树本身对氮需求较大,往往需要施加大量氮肥[5],氮肥虽然对包括杨树等在内的植物的产量和品质起到了一定的促进作用,但是氮肥的大量使用也产生了一系列的生态和环境问题。比如,由于植物吸收氮的能力有限,过量的氮肥得不到利用,会经过挥发、淋洗、硝化等造成损失,产生富营养化,酸雨,温室气体和土壤酸化等环境污染[6]。氮肥过量使用还会造成植物机体氮饱和,而其它营养元素相对短缺,导致植物营养不均衡、生长缓慢和产量降低等负作用[7]。此外,化肥含有多种有毒重金属,过多施用还会引起植物尤其是农产品的污染,引起蔬菜叶硝酸盐含量增加,影响人体健康[8]。因此,有必要提高植物氮利用效率(Nitrogen use efficiency,NUE),使植物在现有氮肥施用状况下,生长表现更好;亦或使植物在降低氮肥施用情况下,生长表现不受影响甚至表现更好,从而推进农林业经济效益和环境友好双赢的发展。
聚天冬氨酸(Polyaspartic acid,PASP)是天然存在于软体动物和蜗牛壳内的一种高分子有机聚合物,是一种对环境友好的、可生物降解的水溶性多肽,其相对分子量为2 000~10 000[9]。PASP 分子含有丰富的酰胺键和羧基等极性基团,赋予其很好的亲水性和水溶性。PASP 可作为土壤保水剂,帮助植物在干旱和半干旱条件下生长[10]。PASP 还具有吸附、螯合和分散等功能[10~12]。对土壤施加PASP 以后,土壤N、P、k、Ca、Mg 等营养元素及B、Mo、Mn 等微量元素的有效性明显提高。此外,PASP 还可以抑制土壤中的铵态氮氮肥的挥发,减少磷肥在土壤中的固定与沉积[13]。PASP 本身虽然不是肥料,但它可以促进植物对养分肥料的吸收,延长肥料利用率来提高植物的养分水平,促进植物生长[9]。研究发现,添加PASP 能够延长肥料释放时间,抑制肥料分解和流失,提高肥料利用效率[14],提高植物根系活力[15~16],改善叶片生理形态[13],增加光合速率[17],调节氮生理代谢相关酶活[17~20],延缓植物衰老[19],促进作物产量[13~17,21~22]。此外,PASP 还有很强的机械强度和粘性,能将沙土凝聚起来,可作为化学固沙剂,提高植物的适应性和成活率,在生态修复中发挥重要作用[23]。然而目前这些研究多以农作物为材料,PASP 作为增效剂对木本植物氮素生理代谢的影响以及作用机制方面的报道却比较少见。为探究PASP 对杨树生长和肥料利用的影响,本研究以“南林895杨”为植物材料,利用Long Ashton(LA)营养液进行水培处理,设置不同浓度氮和PASP 处理,分析不同氮浓度下不同PASP 对杨树生长及氮素利用的调控效应,以期揭示PASP 在杨树氮素利用过程中的作用及相关调控机制,为提高“南林895杨”氮利用效率,实现减氮增效提供理论依据和技术支持。
1 材料与方法
1.1 实验材料与设计
本研究使用杨树杂交无性系“南林895 杨”(Populus deltoides × P. euramericanacv‘Nanlin 895’)组培幼苗为植物材料,选取大小一致的组培苗(约5 cm高)进行营养液水培处理。水培基础营养液为无氮Long Ashton(LA)营养液(pH 为5.8),主要成分包括:0.5mmo·lL-1KCl,0.9 mmo·lL-1CaCl2,0.3 mmo·lL-1MgSO4,0.6mmo·lL-1KH2PO4,42μmo·lL-1K2HPO4,10 μmol·L-1EDTA·Fe·Na,2μmol·L-1Mn-SO4,10μmol·L-1H3BO3,7μmol·L-1Na2MoO4,0.05 μmol·L-1CoSO4,0.2μmol·L-1ZnSO4和0.2μmol·L-1CuSO4。
通过在LA营养液中施加不同浓度氮(硝酸钾作为唯一氮源)和PASP共设立12个梯度,氮浓度分别为无氮(0 mmol·L-1N)、低氮(0.2 mmol·L-1N)、正常氮(2 mmol·L-1N)和高氮(10 mmol·L-1N),PASP浓度分别为0、100 和200 mg·L-1。每隔6 天换1 次营养液。每个处理水平5重复。每株植株供氮量如表1所示。培养条件为:24℃光照/22℃黑暗,光照强度为5 000 lx,光照时间为16 h,湿度为40%~60%。
1.2 材料采集
不同浓度氮和PASP 组合处理两个月后,记录每株植株的株高、根长、地上部分鲜重、根鲜重、叶片数等,根冠比=根鲜重/地上部分鲜重。选择形态学上端完全展开的叶片(从顶端数第3~4 片叶子),记录叶面积。随后,将杨树分为幼叶(YL)、成熟叶(ML)、茎(S)和根(R)4个组织分别采样,记录各组织鲜重,在液氮中冷冻研磨成细粉后,保存于-80℃冰箱备用。
1.3 光合作用的测定
使用Li-6400(Li-cor,USA)便携式光合仪对“南林895杨”水培苗的净光合速率(Pn,μmol CO2·m-2·s-1)、气孔导度(Gs,mol H2O·m-2·s-1)、胞间二氧化碳浓度(Ci,μmol·mol-1)和蒸腾速率(Tr,mmol H2O·m-2·s-1)进行测定。
1.4 氮代谢相关酶活性测定
硝酸还原酶(NR)活性采用磺胺比色法测定[24]。谷氨酰胺合成酶(GS)参照邹琦[25]的方法测定。谷氨酸合成酶(GOGAT)和谷氨酸脱氢酶(GDH)参考Lin等[26]及Gan等[27]的方法测定。天冬氨酸转氨酶(AspAT)参考唐会会等[28]方法测定。
1.5 硝态氮和铵态氮含量
硝态氮含量测定参考Patterson 等[29]的方法。铵态氮含量采用改良的茚三酮比色法[30]。
1.6 叶片氮素利用效率(NUtE)
将杨树叶片在180℃杀青15 min 后,于65℃的烘箱中干燥至恒重,记录干重-鲜重质量百分比,用于计算叶片生物量干重(g)。NUtE 根据Good 等人的公式计算[31]。杨树幼苗成熟叶的氮素利用率(NUtE)=(处理后成熟叶叶片生物量干重(g)-处理前成熟叶叶片生物量干重(g))/植株的总供氮量(g/株))。本实验中,0 N 所有梯度供氮量为0 g,不做计算,0.2 N 为供氮量0.2 mmol·L-1,2 N 为供氮量2 mmol·L-1,10 N为供氮量10 mmol·L-1,。
1.7 数据处理与分析
用Microsoft Excel 2016 对原始数据进行初步汇总和整理,在进行统计分析之前,对数据进行正态性检验。使用IBM SPSS Statistics 21 统计软件进行数据的方差分析和多重比较,采用单因素方差分析检验不同氮浓度和PASP 组合对杨树幼苗生物量、株高和根长等的影响。P值小于0.05 时,认为均值之间差异显著。
2 结果与分析
2.1 不同浓度氮和PASP 组合处理对“南林895杨”幼苗生长发育指标的影响
在持续两个月的处理之后,各个梯度下的杨树水培幼苗表型发生了明显的变化。多重分析表明,除了根鲜重和叶面积,不同浓度氮显著(P<0.01)影响实验期间杨树幼苗大部分生长发育指标。然而,除了根冠比和叶面积,不同浓度PASP几乎没有显著(P<0.05)影响杨树幼苗生长发育。不同的氮和PASP 组合处理对杨树发育指标(除根长外)均有显著影响(P<0.05),其中对根冠比的影响尤为显著(P<0.001)(见表2)。

表2 不同浓度氮和PASP组合处理下“南林895杨”幼苗生长发育指标的多重分析Table 2 Multiple analysis of growth index of poplar hybrid‘Nanlin895’under different N and PASP concentrations
氮本身对杨树生长有显著影响,如表3 所示,随着氮浓度的增加,杨树根冠比降低,杨树叶片数和株高等有明显提高。以无氮为对照,低氮、正常氮和高氮处理下株高增幅分别达到38.56%、43.28%和41.04%,地上部分鲜重增幅分别达到128.67%、260.48%和434.14%,根冠比下降幅度分别达到37.21%、64.51%和56.86%。添加PASP 在无氮和低氮条件下促进了株高和叶片数增加,以及地上部分鲜重积累,但却显著降低了根冠比,以0 N 为对照,0 N+100 mg·L-1PASP、0 N+200 mg·L-1PASP、0.2 N+100 mg·L-1PASP 和0.2 N+200 mg·L-1PASP 植株的根冠比下降幅度分别达到了43.21%、65.25%、55.25%和61.16%。在正常氮供应条件下添加100 mg·L-1PASP增加了植物株高、叶片数、根鲜重和地上鲜重,而在高氮条件下,无论是添加100还是200 mg·L-1PASP均降低。
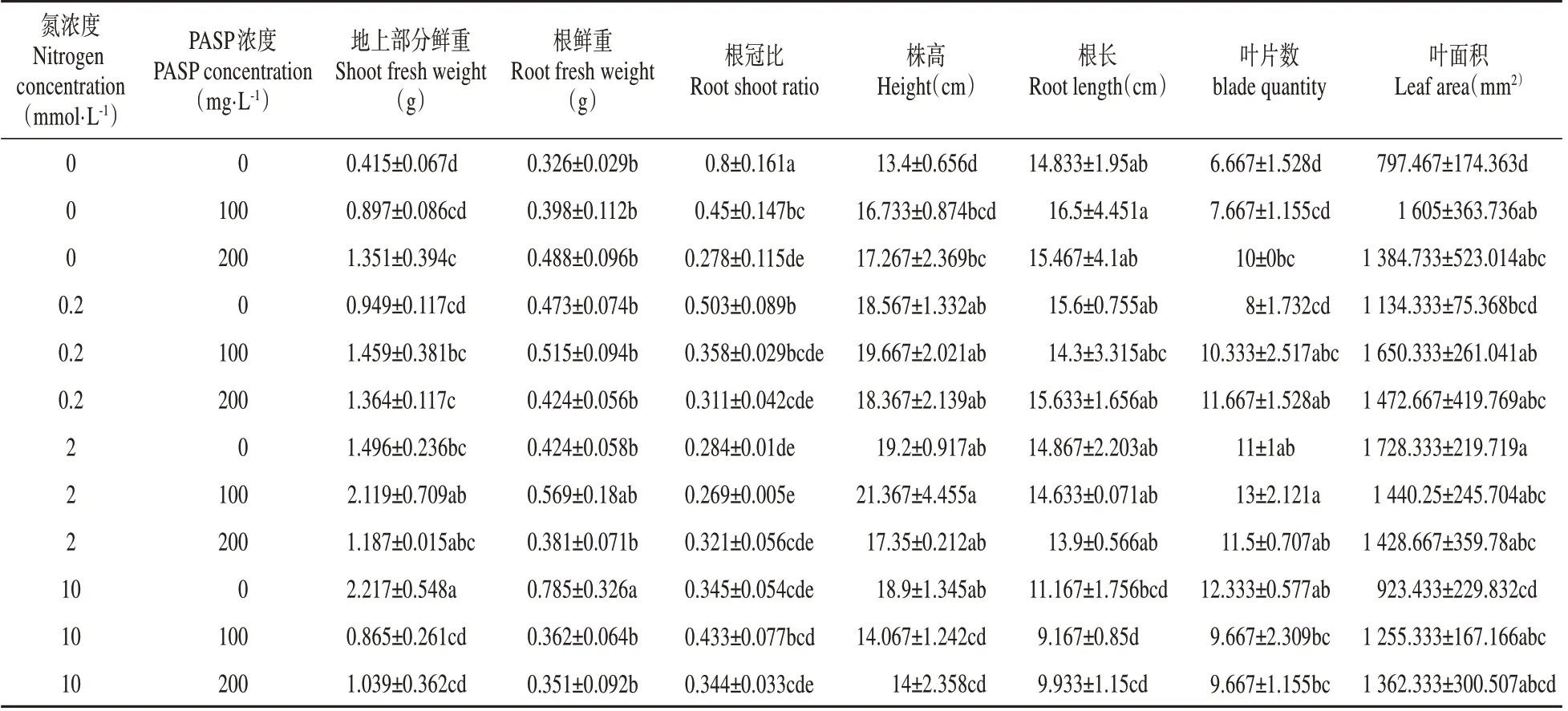
表3 不同浓度氮和PASP组合处理下“南林895杨”的生长发育指标Table 3 Growth index of poplar hybrid‘Nanlin895’under different N and PASP concentrations
2.2 不同浓度氮和PASP 组合处理对“南林895杨”幼苗光合特性的影响
由图1 可以看出,在一定氮浓度下施加PASP明显提高了净光合速率,其中以2 N+100 mg·L-1PASP的净光合速率最高,10 N+100 mg·L-1PASP的净光合速率最低。在无氮条件下施加100 mg·L-1PASP使净光合速率增加了62.27%,施加200 mg·L-1PASP 使净光合速率增加了82.20%;在低氮条件下施加100 和200 mg·L-1PASP 分别使净光合速率增加了27.04%和39.09%;在正常氮条件下施加100 mg·L-1PASP 使净光合速率上升4.64%,施加200 mg·L-1PASP 则使净光合速率降低7.04%;然而高氮的各个处理组净光合速率均较低,施加PASP 加重了对净光合速率的抑制,施加100 和200 mg·L-1PASP 分别使净光合速率下降了37.03% 和35.43%。在无氮条件下施加PASP 使气孔导度增加,添加100 和200 mg·L-1PASP 分别使气孔导度增加144.19%和162.79%;在低氮和正常氮条件下,随着PASP 浓度升高气孔导度先增加后下降,在高氮条件下施加不同浓度PASP 使气孔导度下降,降幅达到25%~31.67%。总体上,胞间二氧化碳浓度在低氮和正常氮处理下保持较低的浓度水平,而在高氮处理下浓度较高,与净光合速率无明显相关性。蒸腾速率的变化与气孔导度正相关。
2.3 不同浓度氮和PASP 组合处理对“南林895杨”幼苗氮代谢相关酶活性的影响
为了进一步研究PASP 影响氮同化作用的生理机制,对氮代谢相关的酶,包括硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)、谷氨酸脱氢酶(GDH)和天冬氨酸转氨酶(AspAT)等的活性进行了测定。多重分析表明,氮浓度极显著(P<0.001)的影响实验中五种酶的活性,NR、GS、GDH 和AspAT 受PASP 影响显著(P<0.05),不同浓度氮和PASP 组合处理显著(P<0.01)影响了NR、GS、GOGAT 和AspAT,而GDH活性没有显著差异(见表4)。

表4 不同浓度氮和PASP 组合处理下“南林895 杨”幼苗氮代谢相关酶活性多重分析Table 4 Multiple analysis of nitrogen metabolism-related enzyme activities of poplar hybrid‘Nanlin 895’under different N and PASP concentrations
由图2 可见,不同浓度氮和PASP 处理对NR酶活性的影响较大。首先,氮浓度对NR 有较大影响,与无氮相比,低氮、正常氮和高氮处理的NR 活性分别上升108.25%,376.52%和55.28%。在无氮和低氮条件下,施加100 mg·L-1PASP使NR活性上升,施加200 mg·L-1PASP 使NR 活性下降;在正常氮和高氮条件下,施加PASP 使NR 活性均降低。GS 活性变化趋势和NR 相反,在无氮和低氮条件下,施加100 mg·L-1PASP 使GS 活性下降,施加200 mg·L-1PASP 使GS 活性上升,在正常氮和高氮条件下,GS 活性随着PASP 浓度升高而上升。GOGAT 活 性 在0.2 N+100 mg·L-1PASP 处 达 到 最 大值,在无氮和高氮条件下随着PASP 浓度升高而上升;在低氮条件下,施加100 mg·L-1PASP 时GOGAT 活性上升,而施加200 mg·L-1PASP 时GOGAT活性下降;正常氮条件下,GOGAT 活性变化正好与低氮的相反。AspAT 活性在无氮和低氮条件下,随着PASP 浓度升高而上升,尤其在无氮条件下增幅明显,添加100 和200 mg·L-1PASP 时AspAT 活性增幅分别达到855.15%和2055.30%;除10 N+200 mg·L-1PASP 相 比10 N 活 性 上 升 了39.44%以外,AspAT 活性在正常氮和高氮条件下随着PASP 浓度升高而下降。GDH 活性在无氮和正常氮条件下,随着PASP 浓度升高而上升,在低氮和高氮条件下,施加100 mg·L-1PASP 使GDH 活性下降,施加200 mg·L-1PASP 使GDH 活性上升。总体上,酶活性大多在无氮和低氮条件下变化较明显。
2.4 不同浓度氮和PASP 组合处理对“南林895杨”幼苗硝态氮和铵态氮含量的影响
在本实验所有处理条件下,施加PASP 几乎使硝态氮含量都降低,特别是在无氮和低氮条件下降低幅度更明显(见图3)。以不添加PASP相应氮浓度作对照,在无氮条件下施加100 和200 mg·L-1PASP 使硝态氮含量分别降低49.19%和56.56%;在低氮条件下分别降低40.19%和30.44%。而铵态氮的变化则不同,铵态氮的含量与供氮量正相关(见图3),以无氮处理组为对照,低氮、正常氮和高氮处理组铵态氮含量增幅分别为60.93%、176.46%和262.17%。施加PASP 在无氮和低氮条件下,对铵态氮含量有一定影响,100 mg·L-1PASP使铵态氮含量升高,而其他条件下PASP 对铵态氮含量影响不显著。
2.5 不同浓度氮和PASP 组合处理对“南林895杨”幼苗成熟叶NUtE的影响
氮素利用效率(NUtE)表示了在不同处理下,植物利用氮素合成生物量的能力。由表5可知,在低氮处理下,施加PASP 明显比未施加PASP 提高了杨树成熟叶NUtE,施加100 和200 mg·L-1PASP使NUtE 分别提高53.7%和43.7%。在正常氮处理下,也有类似效果,施加100 mg·L-1PASP尤为显著(41.64%)。然而在高氮处理下,施加PASP 抑制了杨树利用氮素的能力,使NUtE均下降,施加100和200 mg·L-1PASP 时NUtE 下降幅度分别为60.99%和53.11%。

表5 不同浓度氮和PASP 组合处理下“南林895 杨”成熟叶NUtE的变化Table 5 NUtE of mature leaves in poplar hybrid‘Nanlin 895’under different N and PASP concentrations
3 讨论
氮的可利用性对杨树生长起着决定性的作用[32]。在有限的氮条件下,使杨树能充分利用氮,促进其生长发育,提高杨树胸径和品质,对提高杨树的经济价值和生态效益有重大意义。前人研究认为,PASP 作为肥料增效剂,能够显著促进农作物对氮的吸收和利用[9,14,16~17,33]。本研究也发现,施加PASP 可以有效缓解无氮和低氮对杨树幼苗生长的抑制作用,添加PASP 能明显提高杨树幼苗株高、叶片数、地上部分鲜重和NUtE,降低杨树幼苗根冠比和硝态氮含量。氮供应不足时,杨树地上生物量积累的增加可能与添加PASP 后植物净光合速率增加有关。PASP 在氮供应不足的情况下促进植物生物量积累更明显,这一点与Wang 等人观点是一致的[28]。在高氮条件下,施加PASP 明显降低了杨树生物量、叶片数和根长,根冠比略有升高,这可能与高氮条件下施加PASP 加重了高氮胁迫有关。这些实验结果表明,低氮条件生长的“南林895 杨”通过添加一定量的PASP 有望实现减氮不减产,即减肥增效的目的。本实验条件下,与不加PASP 的氮处理相比,以2 N+100 mg·L-1PASP 处理“南林895 杨”生长表现最好,以0.2 N+100 mg·L-1PASP 处理对“南林895 杨”氮的吸收利用的促进效果最好。
前人研究发现营养液中添加一定量的PASP可以很好的降低水培蔬菜中硝酸盐的含量[16]。本研究也发现,施加PASP 能够降低杨树幼苗硝态氮含量,这在无氮和低氮条件下尤为明显,同期NR和AspAT 酶活性增高,表明PASP 能够促进植物体内硝态氮的同化,增加N 代谢流向天冬氨酸的合成和代谢通路。与此同时,施加PASP 使杨树幼苗体内铵态氮含量增加,同期GS 酶活性降低,GOGAT 酶活性增高,由于营养液中没有铵态氮,因此增加的铵态氮主要由N 在植物体内的代谢产生,表明施加PASP 促进了植物体内N 的同化和转化。此外,本研究中NUtE 随着氮浓度的增加而明显降低,低氮时施加PASP 显著的提高了NUtE,然而高氮时施加PASP 却显著的降低了NUtE。分析原因可能是低氮时,PASP 的施加能够螯合和富集营养液中的氮,因而有利于植物吸收和利用有限的氮源。达到正常供氮水平(2 N)的氮浓度后,过多的氮不能被杨树完全吸收,导致了氮在根部积累,PASP 的施加可能进一步增加了氮的积累,局部过多的氮对植物产生毒害,影响植物对氮的吸收和利用,从而抑制植物生长。
