秋海棠(Begonia grandis)的种内表型多样性
2021-03-02李雁瓷付乃峰孙加芝曹建国田代科
李雁瓷 付乃峰 孙加芝 肖 艳 曹建国 田代科*
(1. 上海师范大学生命科学学院,上海 200234;2. 上海辰山植物园/中国科学院上海辰山植物科学研究中心,上海 201602;3. 上海市资源植物功能基因组学重点实验室,上海 201602)
秋海棠属(BegoniaL.)是植物多样性最丰富的类群之一,为被子植物第六大属,目前已有1 900多个种[1~2]。秋海棠(Begonia grandisDryander)隶属于秋海棠科(Begoniaceae)秋海棠属(Begonia)多年生球茎种类,具有明显的地上茎,冬季地上部分干枯死亡,地下茎休眠。广泛分布于我国南北各地,南起云南屏边,最北可至辽宁凌源,东起浙江宁波,最西可达西藏察隅,也是世界秋海棠科分布最北端、抗寒性最强,分布范围最广之一的种类[3]。该种最早的文献记载可追溯到宋代,其花叶优美、历史悠久、文化深厚,具有较高的观赏、药用和食用价值[4]。
秋海棠的种内多样性十分丰富,不同居群在植株大小、分枝、叶色等某些性状上的变异显著,鉴定困难,从而造成其种内分类比较混乱[5]。以往,基于形态多样性对该种的种内分类没有开展系统的比较分析。种内亚种、变种的发表都是建立在简单的形态观察上,或者仅仅依据几份腊叶标本,因此带有一定的主观性。《中国植物志》[6]及Flora of China(FOC)[7]有关该种的种内单元划分均不太合理,在种内关系的处理上存在分歧,依据现有的性状描述检索无法准确定到亚种和变种,在鉴定实践中缺乏操作性,不利于该种的合理利用和信息交流,因此有待进一步地开展定量分析。而数量分类法能对大量的生物学性状进行全面综合的分析,并对所有的分类性状进行等权处理,再根据性状间的亲缘性和相似性进行归类,从而使植物分类学研究从定性描述水平走向精确、可重复的定量分析水平,摆脱传统分类的主观性[8~9]。本研究以39 个秋海棠居群(包括33 个国产野生居群和6 个日本自然化居群)为研究材料,通过数量性状变异分析、质量性状遗传多样性分析、主成分分析和聚类分析对其33个表型性状进行多样性和数量分类研究,旨在初步探讨秋海棠的主要分类特征,揭示主要性状之间的内在联系及各性状所占有的信息比例,试图为秋海棠的种内单元分类及多样性研究提供客观可行的依据。
1 材料与方法
1.1 试验材料
试验材料取自上海辰山植物园观赏植物资源与种质创新课题组的秋海棠苗圃,以2012~2016年从全国范围内采集和从日本引种的秋海棠居群为观测对象。塑料盆种植(口径18.5 cm,高15.2 cm),每盆种植一株(球)。栽培基质为腐叶土和珍珠岩和树皮按5∶1∶1比例混合,盆底垫上一层陶粒以利排水。浇水频度大体为夏季一天1~2 次,春秋季3~5 天1 次,冬季一周1 次,遵循基质单次浇透、保持润而不干的原则。大棚铺设“寒冷纱”进行遮荫,遮光率约为40%,夏季用风机进行降温(表1,图1)。
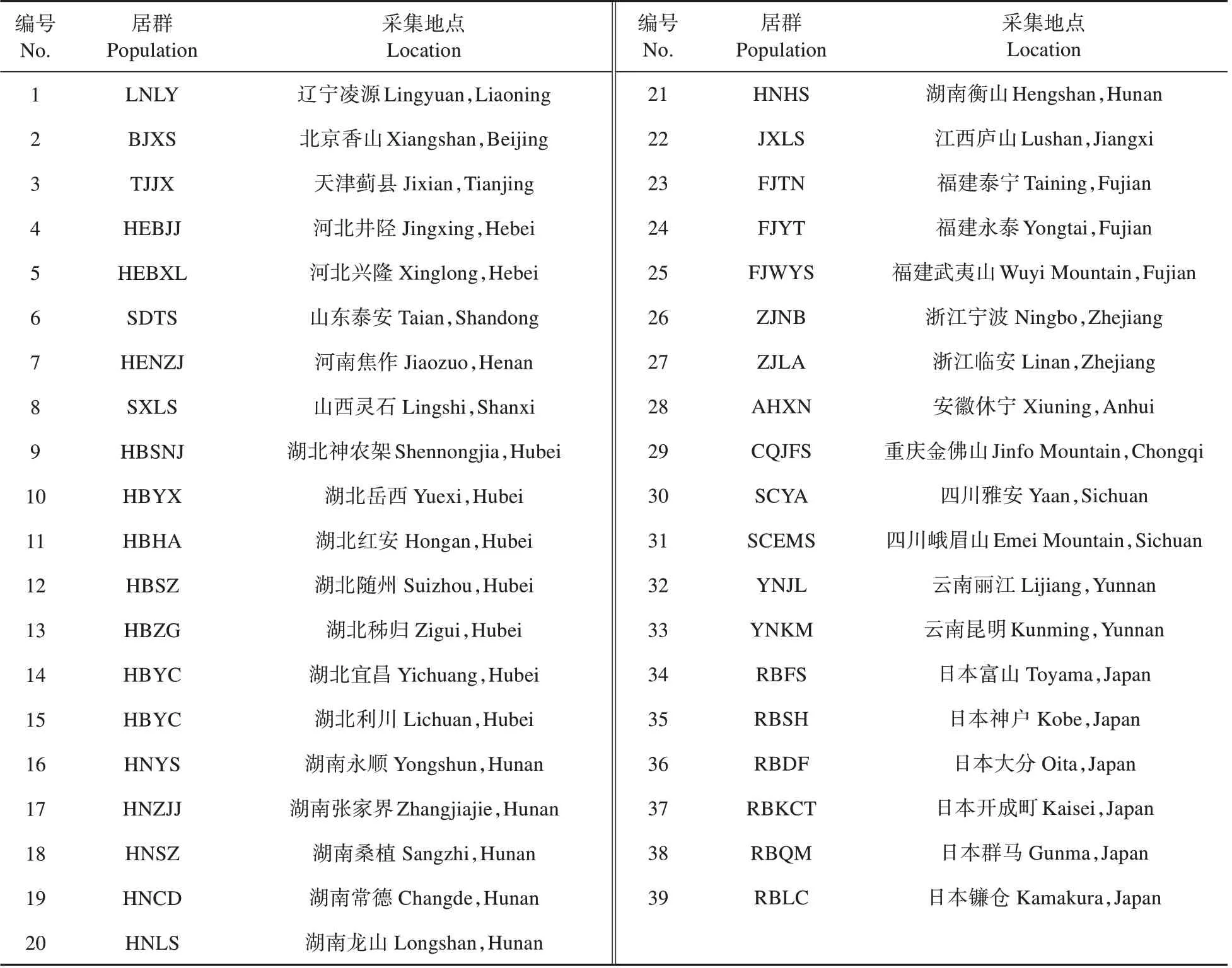
表1 试验材料来源及编码Table1 The origin and code of B.grandis for experiment
1.2 形态观察及数据测量
根据田代科修订的《秋海棠野外性状记录表》(未发表)对苗圃收集栽培的秋海棠植株的完整生长阶段进行观察、记录形态数据和拍照。每个居群选8~10 株健康的植株,于盛花期观测花部等性状,叶片测量从地上茎第一节间开始,从下往上的10 片成熟叶片中随机选取6~8 片进行观测,2018~2019 年连续观察两年。株高、叶柄、叶片、雄花被片等数量性状用直尺测量,每株每个指标测量10次,并充分考虑最大和最小的极端类型,最终取其平均值。其中,叶片测量方法(图2):叶长为中脉轴平行线两端点最长距离;叶宽为同中脉平行线垂直最宽距离;叶宽侧宽为叶片宽侧同中脉平行线垂直最宽处;叶窄侧宽为叶片窄侧同中脉平行线垂直最宽处;下延叶长为从叶柄着生处开始、平行中脉,向下到宽侧叶片底部边缘顶端为止。质量性状采用目测法,结合《中国植物志》[6]和FOC[7]中有关秋海棠的植物资料,对植株进行考察后归纳总结该种在质量性状方面的特征,包括叶斑纹有无、叶脉和叶面颜色、花柄和内外被片颜色等。
1.3 分类性状的选取和编码
对原始数据进行初步统计计算后,将39 个供试居群作为分类单元(Operation taxonomic unit,OUT),以其外部形态为依据,筛选出33 个性状进行后续编码分析(2),其中对立的二元性状(Binary character,简称为“B”)用1 和2 来表示有无;多态性状(Multistate character,简称为“M”)取连续的正整数1、2、3、4、5…进行编码;数量性状(Numerical character,简称为“N”)无需编码,直接取其测量值;导出性状无须编码(Derived character,简称为“D”),取其计算测量值[10-11]。将所有数据测定后录入Excel,汇总为原始数据矩阵。
1.4 数据处理方法
数量性状分析:对于数量性状进行描述性统计,计算其平均值、标准差、极值、极差、变异系数,其中变异系数VC=标准差/均值[12~13]。
质量性状分析:对于二元和多态性状进行形态变异的频数统计,计算遗传多样性指数(Shannon-Wiener diversity index,H′):
式中:pi表示第i种变异类型出现频率;ln为自然对数;H′值可以描述质量性状的总体多样性[14],遗传多样性指数的计算由Microsoft Excel 2016完成[15]。
聚类及主成分分析:使用SPSS 26.0 分析数据,首先对原始数值矩阵进行数据标准化(STD)处理,以消除不同量纲对数据分析产生影响;然后Pearson 相关性为度量指标,采用组间联接对31 个性状进行R 型聚类分析,做出性状指标聚类树形图;同时利用降维对标准化后数值矩阵进行主成分分析,将多项指标转化为少数几项综合指标[16];最后根据R 型聚类分析和主成分分析结果,对居群进行Q 型聚类分析,采用平方欧式距离为度量标准,运用Ward聚类方法做出分类结果树状图。
2 结果与分析
2.1 数量性状的变异分析
对秋海棠的25 个数量性状进行了变异分析,结果见表3。从表3 中可以看出,39 个居群数量性状变异系数为0.077~0.441,变异系数超过0.3的性状有10个,分别为雄蕊群高、叶柄粗、叶窄侧宽、叶宽、雄花内轮被片宽、叶柄长、下延叶长、株高、雄蕊数、雄蕊柱长度,说明这些性状在居群间的差异较大,性状表现不稳定;变异系数低于0.12 的性状有4个,分别为叶片长宽比、叶宽窄侧比、雄花外轮被片长宽比、雄花花冠长宽比,说明秋海棠的叶片形状、外被片及花冠的形状在居群间差异较小,遗传稳定性较高。

表3 秋海棠数量性状变异分析Table 3 Variation analysis for quantitative traits of B.grandis
2.2 质量性状多样性分析
对秋海棠的8 个二元及多态性状进行频率分布和多样性统计,从结果表4 中可以看出:39 个居群数量性状多样性指数变化范围为0.287~1.483。其中,叶斑纹的多样性指数最低,叶片无斑出现频率最高达91.67%,说明叶片出现斑纹的情况较少;地上茎分枝多少、雄花花柄、内外轮被片颜色4 个性状的平均多样性指数均在0.95 以上,说明秋海棠的地上茎分枝及花部颜色在居群间的多样性比较丰富。
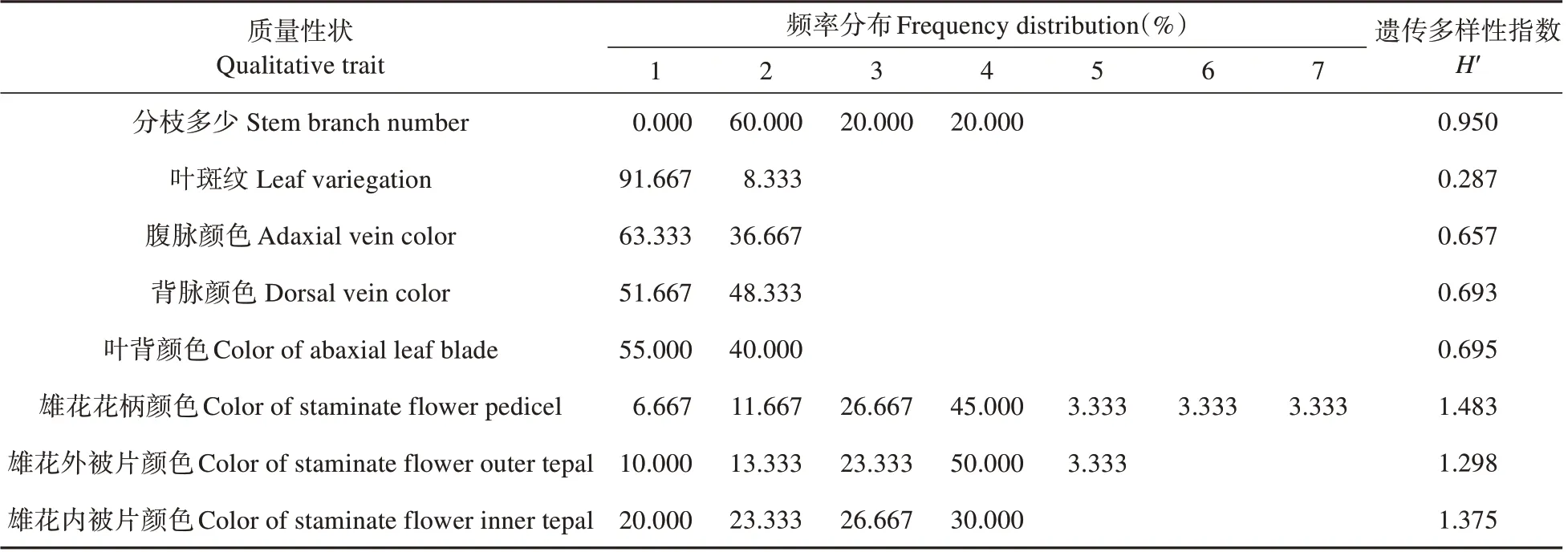
表4 秋海棠质量性状频率分布及多样性Table 4 Frequency distribution and diversity index on qualitative traits of B.grandis

表2 性状及编码Table 2 Taxonomic traits and codes
2.3 主成分分析
主成分分析是将多个性状指标化为少数指标的一种统计方法,能从众多性状因素中找出主要的综合性状因素,使它们尽可能多地保留原始变量的信息,且彼此不相关[17]。对33 个性状进行主成分分析,求得各主成分的特征值、方差贡献率和累计贡献率,结果显示见表5。第1 主成分的特征值为11.29,方差贡献率最大为34.22%,是最重要的主成分,其中,雄花外轮被片长和宽、雄蕊数、雄花花冠长、雄蕊群高和雄蕊群直径的特征向量绝对值较大,说明第1主成分基本反映了这些指标的信息。第2 主成分特征值为4.26,方差贡献率为12.91%,主要由下延叶长、叶宽侧宽、叶长、叶窄侧宽、叶宽等性状决定。第3 主成分特征值为2.85,方差贡献率为8.64%,主要由腹脉颜色、雄花外轮被片颜色、叶脉颜色、叶背颜色等性状决定。前3个主成分累计贡献率为55.77%,是秋海棠分类最主要的区分性状。第4 主成分方差贡献率为6.21%,特征向量绝对值最大的性状为下延叶长同叶长比、雄花内轮被片长宽比和株高。第5主成分贡献率为5.74%,雄花外轮被片长宽比、叶宽窄侧比是特征向量绝对值较大的性状。第6 主成分贡献率为4.57%,特向量绝对值较大的性状为叶长宽比、叶斑纹有无。一般情况下,累计贡献率达到70%以上,事物的基本面貌可以得到反映[17]。前6个主成分的累积方差贡献率为72.28%,包含了39个秋海棠居群的大部分性状信息,贡献率较大的形态学性状是造成秋海棠居群之间表型差异的主要因素,由此可知,秋海棠居群间的差异主要表现为花大小、叶大小、叶色、花色和株高的不同。

表5 主成分的特征值,贡献率及特征向量Table 5 The eigenvalue,contribution rate and eigenvector of selected principal components
2.4 聚类分析
2.4.1 R型聚类分析
数量分类中性状的选取直接影响着分类的结果,因而有必要对所选性状间的关系进行研究[18]。R 型聚类是对研究对象的性状指标进行分类,可以反映出各性状之间的关系及性状选取的合理性[19]。对秋海棠的33 个性状指标进行R 型聚类,结果从树状图(图3)可知,在D=3处做阈值等级分界线,所选性状大多数是独立的,可用于下一步的Q 型聚类分析,但部分性状存在关联性很大的现象。其中,叶长、叶宽、叶宽侧宽、叶窄侧宽和下延叶长这5个性状,雄花花冠宽、雄花内被片长、雄花花冠长、雄花外轮被片长和雄花外轮被片宽这5个性状的相关性较大。因为主成分分析结果表明叶形和花形大小的有关性状是区分秋海棠居群的重要指标,所以都应予以保留。从R 型聚类的结果看,所选的33 个性状可以做到彼此独立影响居群的变异和进化,不会互相造成不必要的干扰。
2.4.2 Q型聚类分析
Q 型聚类是以OTU 为分类基本单位进行的分类运算,可以分析各OUT 间的亲缘关系。对39 个秋海棠居群进行Q 型聚类分析,结果由树状图(图4)可知:在等级结合线L1(D=25)处,39 个秋海棠居群被分为2 大类(图5),第一类包括湖北随州(HBSZ)、山东泰山(SDTS)、湖北宜昌(HBYC)、天津蓟县(TJJX)、湖北秭归(HBZG)、湖南张家界(HNZJJ)、河南焦作(HENJZ2、HENJZ1)、河北兴隆(HBXL)、辽宁凌源(LNLY)、湖北红安(HBHA)、湖北岳西(HBYX)、湖南桑植(HNSZ1、HNSZ2)、湖南龙山(HNLS1、HNLS2、HNLS3)河北井陉(HEBJJ)、山西灵石(SXLS)、湖北神农架(HBSNJ)、北京香山(BJXS)、湖南衡山(HNHS)、浙江宁波(ZJNB)、重庆金佛山(CQJFS)、湖南永顺(HNYS1、HNYS2、HNYS3)21 个居群,主要位于长江以北和华东地区,它们的共同特征是雄花较小,其花冠长宽、内外被片长宽、雄蕊群高与直径、雄蕊数基本上都小于平均值;第二类包括湖南常德(HNCD)、云南昆明(YNKM)、福建永泰(FJYT)、四川雅安(SCYA)、湖北利川(HBLC)、云南丽江(YNLJ)、安徽休宁(AHXN)、福 建 武 夷 山(FJWYS)、福 建 泰 宁(FJTN)、江西庐山(JXLS)、四川峨眉山(SCEMS)、浙江临安(ZJLA1、ZJLA2)、日本富山(RBFS)、日本镰 仓(RBLC1、RBLC2)、日 本 大 分(RBDF1、RBDF2)、日本群马(RBQM)、日本开成町(RBKCT)、日本富山(RBFS)、18 个居群,主要位于西南、华东地区和日本,它们的共同特征是雄花较大,其花冠长宽、内外被片长宽、雄蕊群高与直径、雄蕊数基本上都大于平均值。
在等级结合线L2(D=20)处又可将分出的大类分为4组(图6~7)。第一组包括:湖北随州(HBSZ)、山东泰山(SDTS)、湖北宜昌(HBYC)、天津蓟县(TJJX)、湖 北 秭 归(HBZG)、湖 南 张 家 界(HNZJJ)、河南焦作(HENJZ2、HENJZ1)、河北兴隆(HBXL)、辽宁凌源(LNLY)、湖北红安(HBHA)、湖北岳西(HBYX)、湖南桑植(HNSZ1、HNSZ2)、湖南龙山(HNLS1、HNLS2、HNLS3)河北井陉(HEBJJ)、山西灵石(SXLS)、湖北神农架(HBSNJ)、北京香山(BJXS)、湖南衡山(HNHS)、浙江宁波(ZJNB)、重庆金佛山(CQJFS),主要位于长江以北和华中地区,它们的共同特征是植株较矮、分枝较多、叶片较小,其株高、叶长、叶宽基本上都小于平均值,叶表面没有斑纹,大部分叶片的腹脉、叶背及背脉颜色为绿色。第二组仅湖南永顺的一个特殊居群(HNYS1、HNYS2、HNYS3),位于华中地区,它们的共同特征是植株较高、分枝少、叶片较大,其株高、叶长、叶宽基本上都大于平均值,叶面有白斑,叶片的腹脉、叶背及背脉颜色皆为红色。第三组包括:湖南常德(HNCD)、云南昆明(YNKM)、福建永泰(FJYT)、四川雅安(SCYA)、湖北利川(HBLC)、云南丽江(YNLJ)、安徽休宁(AHXN)、福建武夷山(FJWYS)、福建泰宁(FJTN)、江西庐山(JXLS)、四川峨眉山(SCEMS),主要位于西南和华东地区,它们的共同特征是植株偏矮、分枝较少、叶片偏小,其株高、叶长、叶宽大部分小于平均值,叶表面没有斑纹,叶片的腹脉、叶背及背脉颜色皆为红色。第四组包括:浙江临安(ZJLA1、ZJLA2)、日本富山(RBFS)、日本镰仓RBLC1、RBLC2)、日本大分(RBDF1、RBDF2)、日本群马(RBQM)、日本开成町(RBKCT)、日本富山(RBFS),主要位于华东地区和日本,它们的共同特征是植株较高、分枝较少、叶片较大,其株高、叶长、叶宽基本上都大于平均值,为4组之最,叶表面没有斑纹,大部分叶片的腹脉、叶背及背脉颜色为绿色。
3 讨论
3.1 秋海棠表型性状多样性
变异系数在某种程度上反映出群体数量性状的差异性和多样性,变异系数(CV 值)大则分化程度高、多样性显著,相反则群体相对稳定、一致性较高[20]。选取39 个秋海棠居群的33 个形态学性状,对其中的25个数量性状进行了变异分析,结果显示数量性状变异系数为0.077~0.441,平均变异系数为0.26,表明这些性状在秋海棠居群间变异较大,表型多样性水平较为丰富。同时,对8 个质量性状进行了频率分布和多样性统计,除叶斑纹外,其余7 个质量性状的平均多样性指数均较高(>0.657),表明秋海棠的地上茎分枝、花被片颜色及叶色在居群间的多样性比较丰富。此外,前期性状观测中发现同一居群同时存在叶表面无斑、有白斑,叶背绿色、背脉红色、叶背红色,以及少数开白花个体类型,说明秋海棠的表型在居群内的多样性也很丰富。
3.2 秋海棠分类性状选取
通过对性状指标的主成分分析,共提取了6个主成分。前3 个主成分累计贡献率为55.77%,直到第6 个主成分累计贡献率才达到72.28%,表明各性状的贡献率比较分散,累计贡献率增长比较缓慢,说明在秋海棠的演化过程中性状变异具有多样性[21]。根据各主成分的贡献率选出了影响较大的性状指标,综合而言,花大小、叶大小、叶色、花色、植株大小是秋海棠分类最主要的标准和依据。
R 型聚类分析表明,除叶片和花大小指标有较明显的相关性外,其余性状相对独立,说明性状选取较为合理。但是,由于前期记录的数据不全,雌花(特别是被片数目、柱头形态)和果实(特别是果实及果翅形态)的性状因为数据采集不够而未纳入统计分析,因此,结果无法更全面地揭示各性状间关系。在以后研究中,应进一步补充其他相关重要性状。此外,从湖北宜昌引种的一个居群属于软枝、近匍匐型,花小,综合性状看应独立成枝,也曾被分类学家列为不同变种,但由于栽培管理不当材料死亡,遗憾缺乏相关数据统计,今后有待补充研究。
3.3 秋海棠种内单位划分
秋海棠的种内分类单元的分类学处理一直存在争议。《中国植物志》[6]根据植株高矮、叶片大小及毛被情况、雄蕊柱长度和雄蕊数目、柱头形态、果翅形态等差异将秋海棠处理为1 原亚种、2 亚种、3 变种;FOC[7]根据叶片大小及颜色、雄蕊柱长度雄蕊数目、柱头形态的差异将秋海棠处理为1原亚种、2 亚种;李行娟[22]用三个叶绿体片段联合分析将44 个秋海棠居群划分为三个支系;本研究根据花大小、叶大小、植株大小、叶色、叶斑纹情况将39 个秋海棠居群分为四组,可分成4 个变种。在《中国植物志》和FOC 中,产云南(丽江)的全柱秋海棠(B.grandissubsp.holostylaIrmscher)因花柱分离、不分枝、柱头呈头状或肾状、雄蕊柱长度短于1 mm 的特征区别于其他亚种,而在本研究前期数据统计中,云南丽江(YNLJ)居群雄蕊柱长度有长达3 mm 的,又因花大小、叶片大小、植株大小、叶色等主要性状与其他居群较为相似聚在一起,并没有独立成为一个亚种,同以前的分类相矛盾。研究还发现,日本的几个居群同浙江临安天目山的居群聚类成一枝,说明其亲缘关系最近,结合形态观察和以往的基因大小比较分析,进一步说明,日本的自然化居群的祖先可能来自浙江天目山(田代科此前如此预测过)。
造成以上秋海棠种内单位分类差异的原因有很多方面:一是《中国植物志》和FOC 依据形态学的定性描述对秋海棠进行种内划分,观察样本较少会造成对不重要分类指标的错误侧重,从而导致人为主观设立的性状边界。而数量分类法则是将大量形态学数据进行定量分析,能避免过多次要性状对分类的干扰。二是李行娟所用的叶绿体片段聚类关注的是亲缘关系,而形态学性状的聚类关注的是表型性状,而表现型和基因型之间受到基因表达调控、个体发育调控、生长环境等因素的影响,导致某些表观特征不能完全将遗传变异反映出来[23~24];三是秋海棠居群的性状多样性比较丰富,居群间很可能发生了基因流,存在一些性状过渡类群,因此导致亚种和变种间的界限界定困难,给种内分类带来了挑战。如同一居群的植株叶片颜色有腹脉绿色、腹脉红色和叶背绿色、叶背红色四种个体类型,叶表面有无斑和白斑两种类型,还有一些过渡形态,这说明依据叶色和叶斑纹作为分类标准是不合适的,这同野外观察的实际情况一致。 但是,湖南永顺的一个居群所有个体叶片都具有稳定遗传的白斑,叶片大、植株高但分枝少,即使通过种子播种繁殖的后代也同母本性状一致,性状十分稳定,独立成为一个变种是否比较合理,尚需结合分子证据探讨。综上,《中国植物志》将秋海棠处理为3 个亚种及3 个变种和FOC 秋海棠处理为3 个亚种的处理方式均有待商榷。我们的研究结果和考虑到没有纳入分析的湖北宜昌居群,秋海棠可能分成5个变种更适合。然而,单纯依靠少量形态学性状不能完全反映植物的本质规律,该种居群的种内分类需要综合考虑大量表型性状和分子标记分析结果,今后还需要建立在大量、更全面的采样基础上,结合适当的分子方法开展综合分析才能真正解决问题。
此外,在实际调查过程中,因为多人参与数据记录,发现部分质量性状的记录存在一定的主观性,使得在判断和测量时存在误差。例如叶毛被疏密情况、叶缘齿类型等性状可能存在不同人不同标准的情况,故本次研究没有纳入这些性状指标,但从实际观察来看,这些指标的贡献率不会太高,因此对结果影响不大。在数量分类中,分类性状选取不同会造成不同结果,而本研究没有将雌花和果实纳入分析中,可能影响结果的准确性。因此,今后研究时需要进一步完善形态性状鉴定标准和方法,数据记录应尽可能由一人为主完成,同时使用一些生理指标,如植株萌发出土时间及居群的始花期、盛花期、末花期,并使用多种分类方法进行综合分析以及从群体演化的角度进行系统分类讨论。
