野野村放线菌ATCC 39727利用短链羧酸合成细胞脂肪酸
2021-03-01李志超张广昊
李志超,张广昊,黄 雁,陈 明
(大连工业大学 生物工程学院,辽宁 大连 116034)
0 引 言
脂糖肽抗生素A40926是抗感染新药Dalvance的合成前体[1-2],由野野村放线菌(Nonomuraeasp.)ATCC 39727代谢生成[3-4]。A40926为A0、A1、B0、B1等组分构成的复合物,化学结构上与替考拉宁相似,均含有交联七肽骨架、两个氯原子、一个甘露糖和一个脂酰氨基葡萄糖酸,但各组分连接的疏水脂酰侧链不同[5-6]。研究表明,脂酰侧链能大大提高A40926的抑菌活性[7]。
然而,A40926生物合成基因簇中并不含合成脂酰侧链的基因[8]。Beltramatti和Jovetic等[9-10]研究了培养基中添加支链氨基酸对野野村放线菌细胞脂肪酸和A40926各组分比例的影响,发现A40926各组分比例与生产菌细胞脂肪酸组成在变化趋势上存在一致性,推测细胞脂肪酸经β-氧化后生成A40926的脂酰侧链,而支链氨基酸则作为细胞脂肪酸合成的前体物。
Wallace等[11]研究了3种放线菌细胞直链、支链脂肪酸生物合成的起始物,认为放线菌支链脂肪酸的生物合成以异丁酰CoA(对应生成偶数碳的iso-脂肪酸)、3-甲基丁酰CoA(对应生成奇数碳的iso-脂肪酸)、2-甲基丁酰CoA(对应生成奇数碳的anteiso-脂肪酸)为合成起始物,直链脂肪酸的合成以丁酰CoA(对应生成偶数碳的n-脂肪酸)、丙酰CoA(对应生成奇数碳的n-脂肪酸)为合成起始物。Nonomuraeasp.ATCC 39727作为一种新发现的放线菌,对其生物特性研究较少[12]。本实验对该菌利用短链羧酸合成细胞脂肪酸进行了研究。
1 材料与方法
1.1 材 料
1.1.1 菌 种
Nonomuraeasp.ATCC 39727,实验室保藏。
1.1.2 培养基
培养基ISP2(g/L):葡萄糖4.0,酵母提取物4.0,麦芽提取物10.0,琼脂20.0,pH 7.0。
培养基P150(g/L):葡萄糖20,(NH4)2SO43.65,KH2PO40.3,MgSO4·7H2O 0.45,CaCO35,每升加入3 mL微量元素液(g/L,FeSO4·7H2O 5,CuSO4·5H2O 0.39,ZnSO4·7H2O 0.44,MnSO4·H2O 0.15,CuCl20.02,NaMoO4·2H2O 0.011,50 mL 37% HCl)。
培养基P150中分别添加短链羧酸(丙酸、丁酸、异丁酸、2-甲基丁酸、3-甲基丁酸),质量浓度0.3 g/L。
1.1.3 试剂与仪器
三氟化硼-甲醇,上海安谱科学仪器有限公司;丙酸、丁酸、异丁酸、2-甲基丁酸、3-甲基丁酸、甲醇、正己烷,均为分析纯。
气质联用仪,Agilent Technologies GC-MS 7890/5795。
1.2 方 法
1.2.1 菌体培养
取-80 ℃冰箱中甘油管菌种,涂布至培养基ISP2平板,28 ℃培养7~8 d,挑取单菌落至装有2 mL无菌水的试管中,加入几粒玻璃珠,对菌体进行振荡破碎,取1 mL菌悬液接入25 mL合成培养基中,28 ℃、200 r/min培养72 h。
1.2.2 细胞脂肪酸的甲酯化及组分的GC-MS分析
方法见参考文献[12]。
2 结果与讨论
2.1 培养基P150中菌体细胞脂肪酸组成
菌体经合成培养基P150培养后,对菌体的细胞脂肪酸组成进行GC-MS分析,图1为培养基P150培养菌体细胞脂肪酸的总离子色谱图。结果表明,共检出9种主要脂肪酸,以偶数碳直链脂肪酸为主,C16:0脂肪酸质量分数最高(22.08%),C14:0、C16:1、C16:0、C18:0 4种偶数碳直链脂肪酸质量分数占总脂肪酸的47.76%;偶数碳支链脂肪酸i-C16:0质量分数达21.89%,i-C14:0和i-C16:0两种脂肪酸质量分数达27.04%;奇数碳的i-C15:0、C15:0、a-C17:0脂肪酸同时被检出,质量分数见表1。
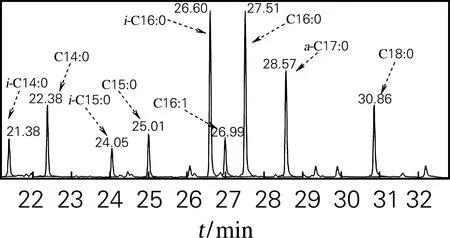
图1 培养基P150中细胞脂肪酸总离子色谱图Fig.1 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150
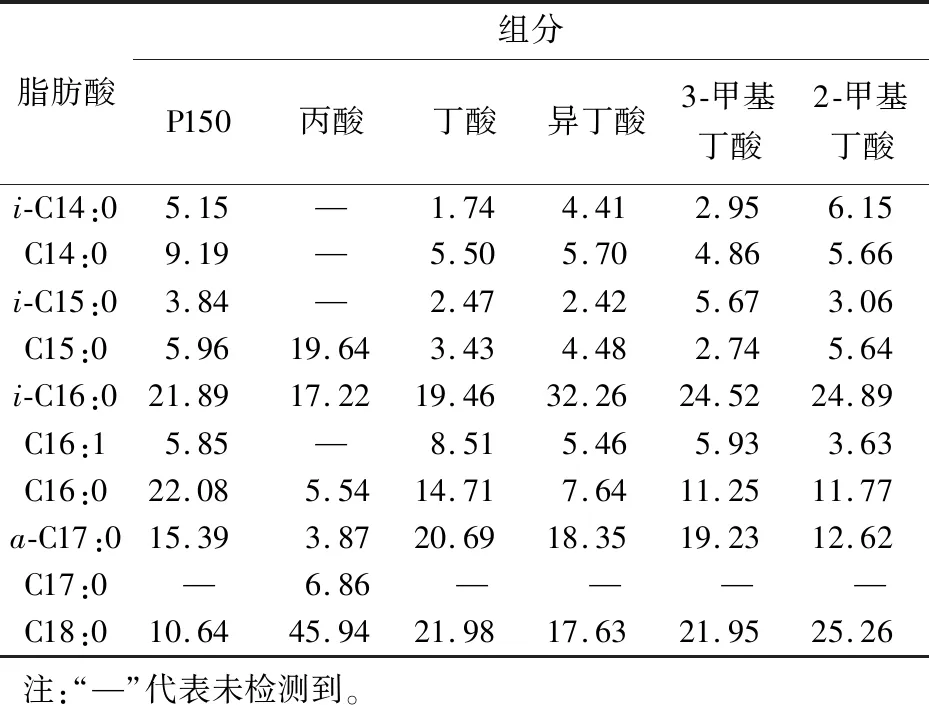
表1 菌体细胞脂肪酸组成Tab.1 Cell fatty acids composition of Nonomuraea sp.ATCC 39727 %
2.2 添加丙酸对菌体细胞脂肪酸组成的影响
图2为合成培养基P150添加0.3 g/L丙酸后菌体细胞脂肪酸的总离子色谱图。与图1相比,添加丙酸后细胞脂肪酸组成发生了很大变化,奇数碳直链脂肪酸质量分数显著增加,其中C15:0 脂肪酸质量分数由原来的5.96%增加到19.64%,原来未检出C17:0脂肪酸,后达6.86%(表1),这可能是丙酰CoA为奇数碳直链脂肪酸合成起始物所致;添加丙酸后,C14:0、C16:1、C16:0等偶数碳直链脂肪酸及anteiso-C17:0脂肪酸质量分数显著降低,但C18:0脂肪酸质量分数大幅度增加。
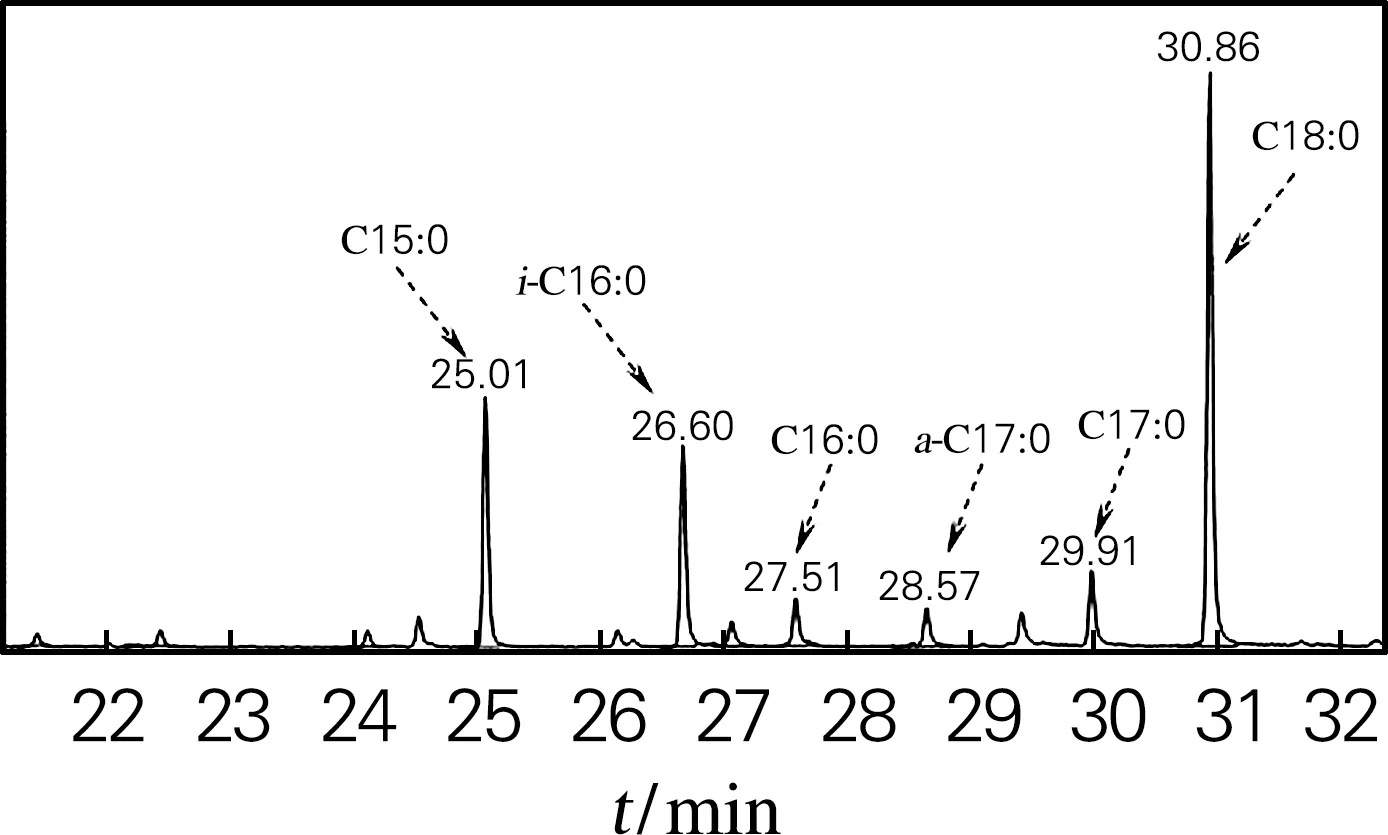
图2 添加丙酸菌体的脂肪酸总离子色谱图Fig.2 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150 supplemented with propionic acid
2.3 添加丁酸对菌体细胞脂肪酸组成的影响
图3为合成培养基P150添加0.3 g/L丁酸后菌体细胞脂肪酸的总离子色谱图。与图1相比,添加丁酸后细胞脂肪酸组成变化不大,添加丁酸后,C16:0脂肪酸质量分数下降,C18:0脂肪酸质量分数增加,C14:0、C16:1、C16:0、C18:0等4种偶数碳直链脂肪酸质量分数占总脂肪酸质量分数的50.70%,质量分数变化不大(见表1),说明添加丁酸对偶数碳直链脂肪酸的合成起始物丁酰CoA质量分数的影响并不大。

图3 添加丁酸菌体脂肪酸总离子色谱图Fig.3 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150 supplemented with butyric acid
2.4 添加异丁酸对菌体细胞脂肪酸组成的影响
图4为合成培养基P150添加0.3 g/L异丁酸后菌体细胞脂肪酸的总离子色谱图。与图1相比,添加异丁酸后细胞脂肪酸组成发生了很大变化,偶数碳支链脂肪酸i-C16:0质量分数显著增加,为质量分数最高的脂肪酸,同时C16:0脂肪酸质量分数显著下降(表1)。异丁酰CoA为i-C16:0 脂肪酸合成起始物,异丁酸的添加导致了i-C16:0脂肪酸质量分数的显著增加,同时导致偶数碳直链脂肪酸C16:0质量分数显著降低。

图4 添加异丁酸菌体脂肪酸总离子色谱图Fig.4 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150 supplemented with isobutyric acid
2.5 添加3-甲基丁酸对菌体细胞脂肪酸组成的影响
图5为合成培养基P150添加0.3 g/L 3-甲基丁酸后菌体细胞脂肪酸的总离子色谱图。与图1相比,添加3-甲基丁酸后细胞脂肪酸组成变化不大。3-甲基丁酰CoA为奇数碳iso-脂肪酸的合成起始物,添加3-甲基丁酸后i-C15:0脂肪酸质量分数增幅很小(表1),推测是Nonomuraeasp.ATCC 39727利用3-甲基丁酰CoA为脂肪酸合成起始物的能力较差所致。
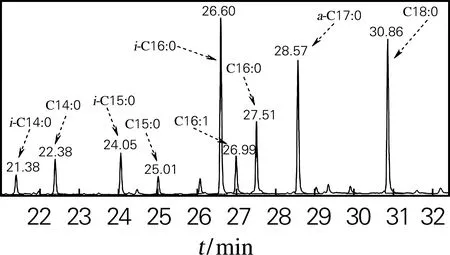
图5 添加3-甲基丁酸菌体脂肪酸总离子色谱图Fig.5 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150 supplemented with 3-methylbutyric acid
2.6 添加2-甲基丁酸对菌体细胞脂肪酸组成的影响
图6为合成培养基P150添加0.3 g/L 2-甲基丁酸后菌体细胞脂肪酸的总离子色谱图。2-甲基丁酰CoA为奇数碳anteiso-脂肪酸的合成起始物,但添加2-甲基丁酸却导致a-C17:0脂肪酸质量分数有所降低(表1),因此Nonomuraeasp.ATCC 39727体内anteiso-脂肪酸的合成还需要进一步的研究。

图6 添加2-甲基丁酸菌体脂肪酸总离子色谱图Fig.6 FAME GC-MS profile of Nonomuraea sp.ATCC 39727 cells grown in medium P150 supplemented with 2-methylbutyric acid
3 结 论
对脂糖肽抗生素A40926产生菌Nonomuraeasp.ATCC 39727利用短链羧酸合成细胞脂肪酸的能力进行了研究。该菌优先利用异丁酰CoA、丁酰CoA和丙酰CoA为细胞脂肪酸合成起始物,但利用3-甲基丁酰CoA为脂肪酸合成起始物的能力较差。
