沙生植物沙鞭不同居群形态变异研究
2021-02-26梁瑞芳刘玉萍
吕 婷 刘 涛 梁瑞芳 苏 旭,3 刘玉萍*
(1. 青海师范大学地理科学学院,西宁 810008;2. 青海师范大学青海省青藏高原药用动植物资源重点实验室,西宁 810008;3. 青海师范大学生命科学学院,西宁 810008)
沙鞭(Psammochloa villosa Bor.)是禾本科(Po⁃aceae)沙鞭属(Psammochloa)的一种多年生根茎型草本植物,主要分布于我国西北部,常生长在流动和半流动沙丘,以及沙地半灌木丛中[1~3]。与其他沙生、旱生植物一样,沙鞭因长期适应干旱、沙埋等环境胁迫而形成了典型的旱生沙生特征,对流动沙丘具有很强的适应性,属于草原带、荒漠草原带以及荒漠地区流动沙丘上分布的沙地植物群落的优势种[3~7]。沙鞭作为一种典型的固沙植物,可以通过根状茎进行快速的克隆生长,不仅具有重要的生态价值,而且也因花序相对粗长、穗多粒大等优点是农牧业上禾草良种繁育和牧草利用的重要基因资源[8]。由于沙鞭分布环境的多样性,导致不同居群之间在植株高度、叶和花等各方面都存在较大差异。截至目前,国内学者对沙鞭种质资源的研究主要集中在克隆整合特性、克隆生长特性、分株种群特征、种子生物学以及遗传多样性等方面,譬如,董鸣[9~10]通过切断根茎对根茎禾草沙鞭和赖草克隆生长影响的比较研究发现,沙鞭的克隆整合特性和克隆生长特性使它在水分短缺、营养贫瘠、生境斑块化和经常遭受扰动的沙地环境中具有较强的生存能力;黄振英[3]通过对沙鞭种子在流动沙丘上传播、萌发、幼苗出土和存活等一系列的研究,旨在探讨沙鞭种子在成熟后的生理生态状态,以及生境条件对萌发的影响;Li[11]利用ISSR 分子标记技术对分布在我国西北地区的7 个沙鞭种群的遗传变异和克隆多样性进行了研究,结果发现种群间的遗传变异高于种群内的遗传变异,且利用ISSR 分子标记技术研究沙鞭遗传多样性的效率远高于等位酶电泳技术。然而,目前针对沙鞭种群遗传变异的研究仅见于分子标记技术和等位酶电泳技术的研究,而基于表型性状对沙鞭遗传变异的研究尚属空白。鉴于此,本研究对分布于内蒙古高原及毗邻地区沙鞭不同居群的形态特征进行观测,旨在揭示沙鞭不同居群间表型形态性状变异的特点及其与地理环境的关系,为进一步研究沙鞭种质资源的保护、开发利用和优良品种的筛选提供理论依据。
1 材料和方法
1.1 实验材料
2016 年7~8 月,对分布于我国典型区域的沙鞭种质资源的生境特点和生态特性进行了观测和样品收集,需选择生长正常,无严重缺陷,无明显病虫害的植株。本文随机选取20个具有代表性的居群测量其表型性状,每个居群取5~10 株个体,个体间水平距离应保持在200 m以上,以避免采集到同一植株的亲缘样本。同时,采用GPS 定位系统记录其生长地、经度、纬度以及海拔等基本信息,各居群的地理位置概况详见表1。
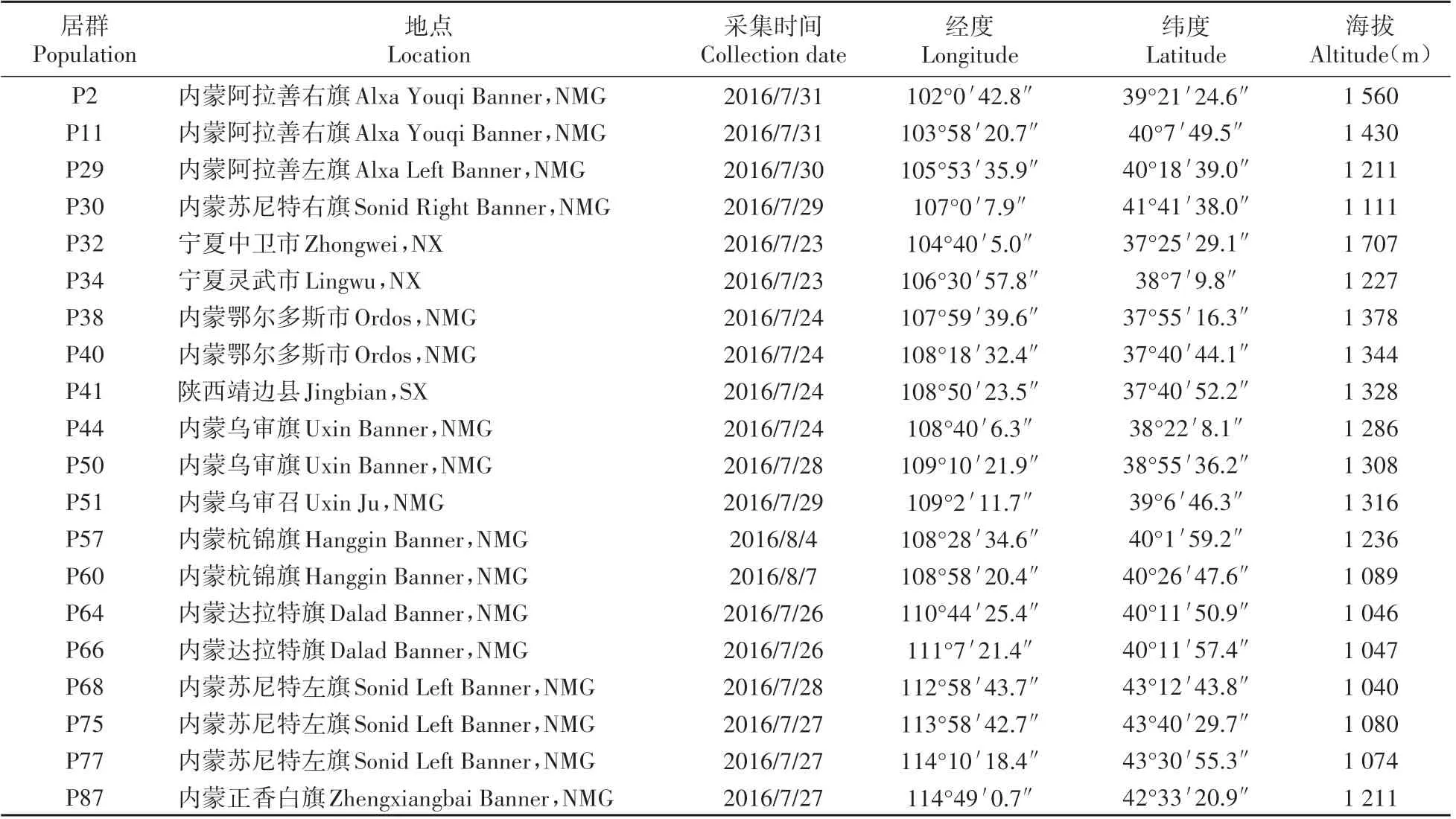
表1 各居群的地理位置与生境概况Table 1 Location and the habitat characters of populations
1.2 形态性状的观察和测量
利用直尺、游标卡尺等测量工具对沙鞭不同居群的12个数量性状值进行测量,具体包括株高、花序长度、旗叶长度、旗叶宽度、旗叶长宽比、小穗长度、外稃长度、第一颖长、第二颖长、颖脉数、外稃脉数和内稃长度。其中,株高是从植株根颈部到顶部之间的距离。每个居群选取6株,个别居群因标本数量较少而选取2~5 株进行观测[8],将所测的各居群的表型性状值录成形态数据表。
1.3 数据处理
利用SPSS 16.0 软件进行统计分析,计算各表型性状的极差、最小值、最大值、标准差和变异系数,并进行单因素方差分析、聚类分析和主成分分析。其中,对居群内各形态特征变异度的测度以变异系数为基准;用单因素方差分析(ANOVA)表示居群间形态变异度;以聚类分析来衡量居群间的形态分化,并绘制分类树状图;用主成分分析(PCA)确定哪些性状是造成沙鞭表型差异的主要因素[12]。
2 结果分析
2.1 表型性状基本统计分析
本文对采集到的109 份沙鞭标本的12 个表型性状进行统计分析表明(见表2),各性状存在不同程度的变异。从总体上看,变异系数在9%~30%,平均变异系数为18.46%,其中,旗叶长宽比的变异系数最高,其变异系数为29.66%,变异幅度为21.16~104.20 cm;小穗长度的变异系数最低为9.73%,变异幅度为1~2 cm。变异系数反映的是性状值离散程度大小,当变异系数越大,则性状值离散程度越大[13~14],而变异系数从小到大依次是:小穗长度<内稃长度<第二颖长<外稃长度<第一颖长<株高<外稃脉数<花序长度<旗叶长度<旗叶宽度<颖脉数<旗叶长宽比,由此说明,相比于其他性状,旗叶长度和旗叶宽度的稳定性更高。另外,一般认为当样本间存在显著差异时其变异系数大于10%[15]。因此,除了小穗长度和内稃长度的变异系数低于10%,其他性状的变异系数均大于10%,表明沙鞭大部分表型性状表现出丰富的多样性。
2.2 形态学性状值差异显著分析
通过对12个具有代表性的表型性状进行F检验(见表3),其中颖脉数和旗叶长宽比达到显著水平(P0.05<P<P0.01);而其余的表型性状均达到了极显著水平(P<P0.01)。从总体上看,各表型性状的自由度均相等,F 检验值范围为1.832~8.958。其中,株高的F 检验值最大,而旗叶长宽比最小。在P<0.05时,各性状的居群间方差分量均大于居群内方差分量。同时,利用ANOSIM方差分析探讨居群内和居群间的差异显著性,当R=0 时,说明居群间没有差异;当R>0 时,表示居群间差异大于居群内差异。从图1 中可以看出,R=0.776(P<0.05),由此说明沙鞭的各性状变异均为居群间大于居群内。
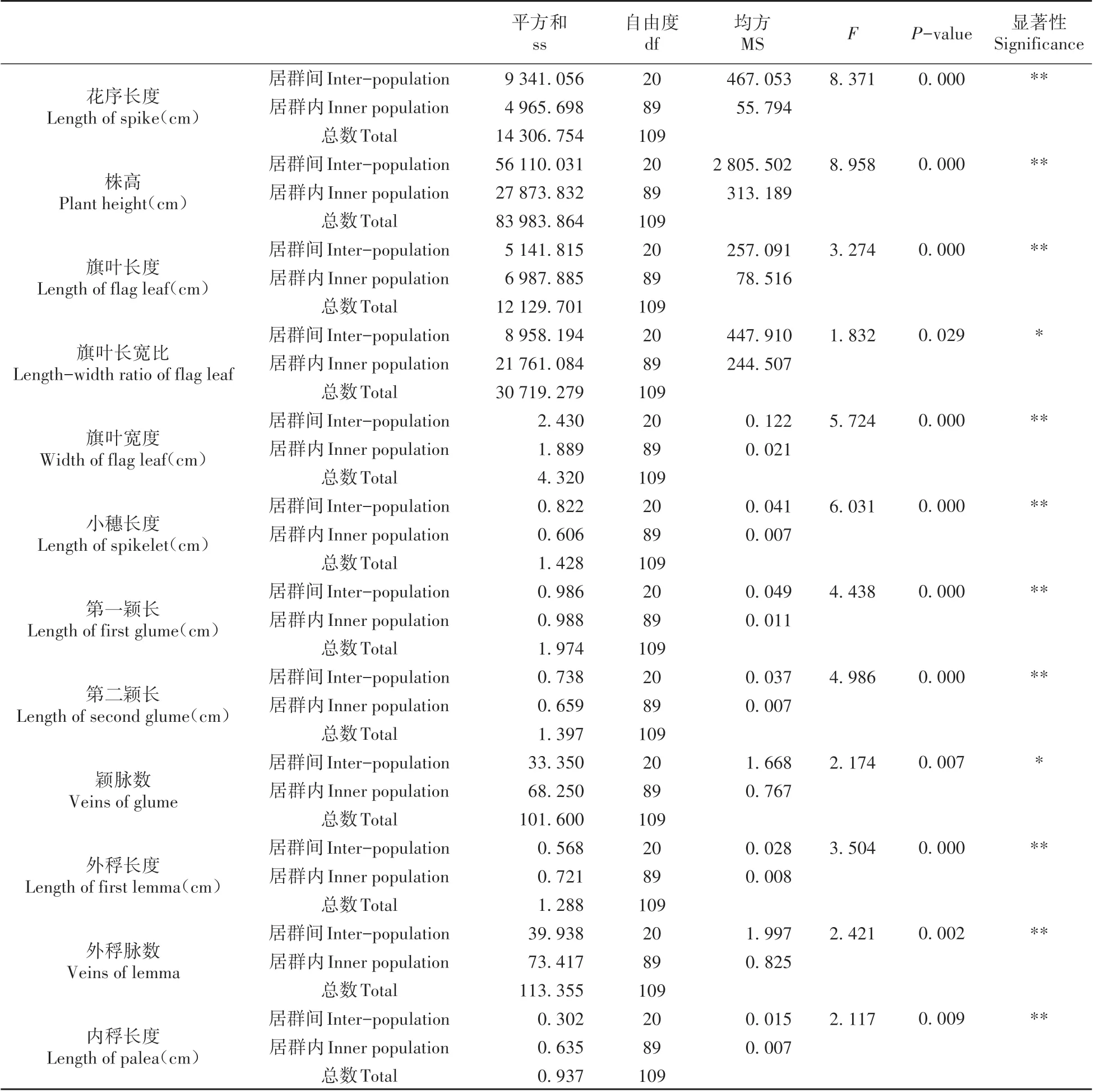
表3 性状值差异显著性分析Table 3 Difference Significance Analysis of Character Values
2.3 表型聚类分析
利用UPGMA 法对20 个自然居群进行聚类分析,从分类树状图中可以看出(见图2),12 个表型性状在不同居群间表型形态有较大的差异,在欧式距离为20.5 时,可将参试居群分为两大类群,而这些居群均选自不同地理位置,具有一定的代表性,但沙鞭居群各表型性状的聚类划分并没有依据地理距离而聚类,由此说明沙鞭群体间表型性状变异的不连续性。
2.4 主成分分析
沙鞭不同居群间均存在丰富的遗传变异,为探讨引起变异的主要原因,故通过主成分分析来显示各性状在表型多样性构成中的作用[16]。主成分分析(Principal Component Analysis)是一种利用降维的思想将多指标转化为少数几个综合指标,使这几个少数综合指标可以反映原来所有指标的信息的多元统计方法[17]。主成分分析结果表明,当m=4 时,累计贡献率为82.277%,说明前4 个主成分可近似代表原始因子的大部分信息(见表4)。其中,第一主成分占40.399%,载荷最高的是第二颖长,其次是小穗长度、第一颖长、内稃长度、株高、外稃长度和花序长度,反映了生殖器官的特点;第二主成分占19.261%,对其影响最大的是旗叶宽度,其次为旗叶长度,反映了营养器官的特点;第三主成分占12.538%,对其作用最大的是旗叶长宽比,其次是外稃脉数,主要反映营养器官的特点;第四主成分占10.079%,主要形状为颖脉数,反映了生殖器官的特点。综上所述,在前4个主成分中旗叶长度、旗叶宽度、颖片长度、小穗长度等是造成表型多样性的主要因素。通过提取分析前3个主成分所绘制的三维空间分布图(见图3)与聚类分析结果基本一致。
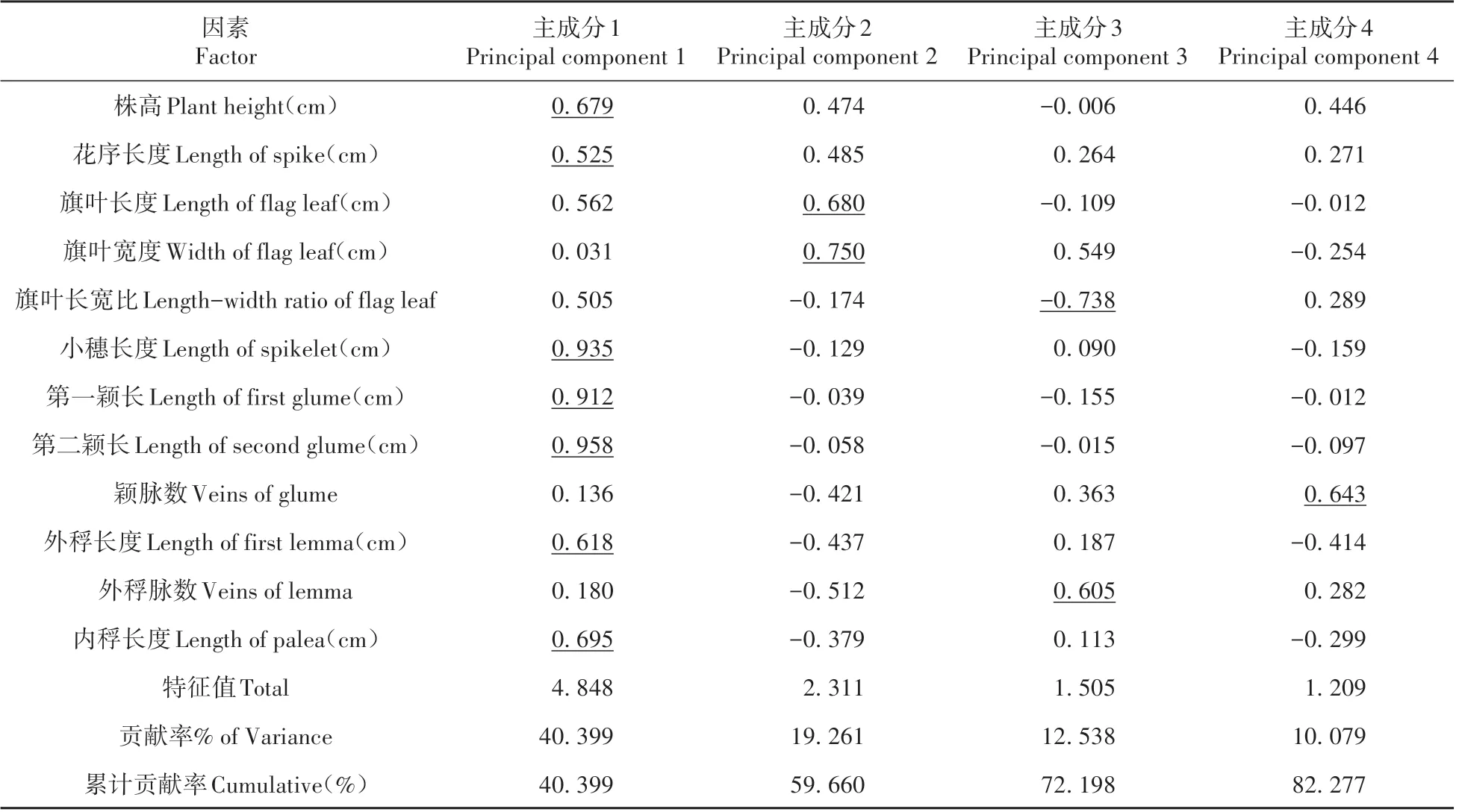
表4 沙鞭种植资源的主成分分析Table 4 Principal component analysis of germplasm resources of P.villosa
3 讨论
在生物学中,变异是指种与个体之间的各种差异,是生物进化和人类育种的根源,包括种群的个体表型变异、环境变异和遗传变异。其中,表型变异是以植物的可塑性为基础的,表现在基因型相同的个体因外部环境条件状况不同而形成不同的表型[17]。那么,种下变异作为居群和个体在时空中发展、变化的产物,是物种形成与进化的基础[17]。通过对居群内和居群间的变异式样和变异规律进行深入探讨,旨在为更加全面准确地理解种内个体所表现出的形态差异,进而揭示物种形成与进化的机制提供帮助[18],同时还可为种下等级的划分和种的界定提供依据。因此,本文通过对参试的109 份沙鞭种质资源的12 个表型性状的多样性进行分析,结果表明除小穗长度和外稃长度外,其余各表型性状的变异系数均在10%以上,即沙鞭的表型性状在各居群间和居群内均存在丰富的变异,且不同表型性状的变异幅度各不相同。同时,不同居群间表型性状的变异数据显示:叶部表型性状(3项)的平均差异系数比穗部性状(7个)的大,由此说明生殖器官性状指标的稳定性大于营养器官。另外,通过主成分分析发现沙鞭种质资源12 个表型性状可以转化为4 个主成分,且代表了表型性状82.277%的信息,而造成表型多样性的主要性状包括旗叶长度、旗叶宽度、颖片长度、小穗长度等。因此,在对沙鞭种质资源进行初步筛选时可以把上述性状作为评价标准[19]。此外,沙鞭表型性状在居群内和居群间均存在明显的遗传变异,但以居群间变异为主,而引起居群间表型性状的变化原因是多样的,其一,从地理环境角度而言,内蒙古高原年均温分布呈西高东低,年降水量分布呈东多西少,由于不同地理位置选择压力的不同,以及地理分布呈间断的零散分布造成基因交流不频繁,使得居群间的表型变异较大[16]。其二,从生殖方式角度而言,沙鞭种质资源既能克隆繁殖,也能种子繁殖。但在自然条件下,一方面由于营养枝生长旺盛,而生殖枝常受到抑制,故不能每年结实[20];另一方面,当光照、花期不遇时会导致结实少或成熟难。因此,通过鸟类或动物取食传播较困难[16],故常以根状茎繁衍出分株。以上可以看出沙鞭表型性状的变异不仅与遗传因素有关,也与环境饰变的影响密不可分[17,21]。最后,从聚类结果来看,表型性状的聚类与地理来源没有明显的相关性,且不同分布区域的居群其表型形态出现了交叉遗传,引起这种现象的原因可能是受环境因素、人为因素和其他因素(天气等)的影响[22]。
鉴于此,制定有效的资源收集保存和育种策略具有重要的现实意义。在选育新品种时,一方面选择具有优良性状的植株,如植株高大,单株叶面积大,果穗较多的株系;另一方面选择表型变异大的亲本杂交,可以栽培出更多不同类型的优良个体[23]。除此之外,在选择育种亲本时还应扩大物种的采集范围,寻找更有价值的材料。另外,沙鞭表型变异丰富,有利于种群的稳定,故应对优良植株的收集、筛选、保存和培育加大力度,旨在保持和提高沙鞭种质资源的多样性和丰富度,并为沙鞭种质资源的进一步收集、保存和遗传改良提供科学依据。
