梓属5个种对干旱胁迫的生理响应
2021-02-26于晓池杨桂娟董菊兰王军辉麻文俊
于晓池 杨桂娟 董菊兰 王军辉 麻文俊 张 鹏
(1. 东北林业大学,哈尔滨 150040;2. 中国林业科学研究院林业研究所,林木遗传育种国家重点实验室,国家林业和草原局林木培育重点实验室,北京 100091;3. 甘肃省小陇山林业实验局林业科学研究所,天水 741022)
干旱已经成为最常见的非生物胁迫之一,对林业发展造成的危害日趋严重[1~2]。为了缓解这一环境问题对林业生产的影响,不仅可以实施人工水肥管理[3~4]、树体修枝管理[5]、林分结构调控管理[6~7]等技术措施,也可以通过抗旱性优良新品种选育[8],提高林木的抗旱能力。
叶片是植物与外界环境进行物质与能量交换的主要器官,对环境变化较为敏感[9]。环境会造成叶片形态与生理方面的改变,而叶片结构的变化也会对植物应对外界环境的能力造成影响[10]。干旱胁迫使叶片积累大量活性氧,对叶片造成损伤,还会降低叶绿素含量、引起叶片荧光参数的变化,同时,干旱还会使气孔关闭,最终导致光合作用的减弱[11~15]。赵燕和王丁等人则认为,干旱会降低叶水势[17],而叶水势的降低可以调节幼苗与土壤水势之间的梯度,从而增加植物的吸水能力[18~19]。因此叶片已经成为研究植物对逆境适应机理的重要方向[14]。
梓属(Catalpa Scop.)为紫葳科(Bignoniaceae)乔木,全世界约13 种,分布于美洲及亚洲东部,我国 引 入 种 共5 种 及1 变 型[20]。包 括 楸 树(C.bungei)、灰楸(C.fargesii)、黄金树(C.speciosa)、梓树(C.ovata)、藏楸(C.tibetica)和滇楸(C.fargesii f.duclouxii)、其中滇楸为灰楸的变形,各树种材质良好,为珍贵用材树种。从形态结构、亲缘关系和开花结果规律分析,可将6个种分为楸树组和梓树组2组,楸树、灰楸和滇楸属于楸树组。臧楸、梓树和黄金树属于梓树组[21]。本文通过人工模拟梯度干旱胁迫处理,分析了梓属5 个种:黄金树、梓树、灰楸、滇楸和楸树的叶片性能,包括叶片水势(ΨL)、气孔导度(Gs)、叶绿素相对含量(SPAD)、叶绿素荧光参数和比叶重(LMA)对干旱胁迫响应,为梓属耐旱树种的筛选、抗旱育种及栽培工作提供理论依据。
1 试验地概况
试验设在甘肃省天水市小陇山林业科学研究所温室内,位于34°28′N,105°54′E,海拔高度1 160 m,气候属暖温带半湿润型,年降雨量600~800 mm,年蒸发量1 290.0 mm,年平均气温10.7℃,≥10℃积温3 359.0℃,极端高温39℃,极端低温-19.2℃。正常年份4~9 月份晴天日平均光照时数13~14 d,夏季日最高光照强度2 500 μmol·m-2·s-1,无霜期约190 d。温室面积为400 m2,高3 m,温室顶部覆盖材料为PC 透光板,夏季温室内部光强可达800 μmol·m-2·s-1,温室四周围墙高1.2 m,为使温室内部通风透气良好,围墙以上至顶部未加装隔离材料,温室内夏季温度最高为39℃。
2 材料与方法
2.1 试验材料
试验选取梓树属的黄金树、梓树、灰楸、滇楸和楸树为试材,于2008 年3 月将苗木栽植在塑料盆内,盆高25 cm,内径23 cm,苗木培养土为黄绵土,田间持水量为29%,放置在小陇山林科所玻璃温室内,每个种栽植20株苗木。
2.2 试验设计
从3 月开始将种子苗置于充足水分条件下生长,每天早上8:00~9:00 浇水,以苗木花盆底座湿润为止。到6 月中旬苗木高达到25 cm 以上时开始控水试验。将试验苗木在2 种水分条件下培养50 天。每种每处理重复5 株,每处理按完全随机区组排列。2种水分处理每次的浇水量为:700 mL(正常水分条件)、400 mL(干旱胁迫),每2 d 浇一次水,浇水时间为18:00点。
2.3 测定指标
8 月中旬测量苗高(cm)和地径(cm),用扫描法测定叶面积(cm2);采用SC-1气孔计在测定气孔导度(Gs);用SPAD 叶绿素计测定叶绿素相对含量(SPAD);用OS-30P 叶绿素荧光仪在凌晨4:00~5:00 测定叶绿素荧光参数;用WP4 水势仪在凌晨4:00~5:00测定叶片水势(ΨL);后对叶片进行烘干称重并计算比叶重(LMA),计算公式为:
比叶重(LMA)=叶片干重(g)/叶片面积(cm2)(1)
3 数据处理
Excel 2003 进行数据整理和分析;SPSS16.0 进行单因素(one-way ANOVA)、双因素(two-way ANOVA)方差分析,检验不同树种各性状是否存在显著差异,多重比较采用Duncan 法;Graphpad Prism 7进行作图。
采用隶属函数法评价梓属各树种的抗旱能力差异。计算各指标的隶属函数值。当指标与抗旱性呈正相关,则用公式(2);当指标与抗旱性呈负相关,则用公式(3)。综合评价值用公式(4)~(6)计算。
式中:μ(Xij)为i 树种j 指标的隶属函数值,Xij为i 树种j 指标的测定值,Xjmin为各树种j 指标测定最小值;Xjmax为该指标测定的最大值。
式中:Vj为标准差系数,Wj为各指标的权重,µ(Xj)为树种j指数的隶属函数值,D为综合价值。
最终根据D 值对各树种进行抗旱综合性评价。
4 结果与分析
4.1 干旱胁迫对生长的影响
经过50 d 的两种水分处理,在同一水分条件下树种之间的苗高(P<0.01)和地径(P<0.01)差异极显著(见图1)。在2 种水分条件下,楸树的苗高均最高,其次为滇楸,黄金树和灰楸,梓树的苗高最低。而地径则是楸树和黄金树较高,滇楸,灰楸和梓树较低。在700 mL 和400 mL 水分处理下,苗高极差分别为:20.7和26.2。说明干旱胁迫增加了各个种的苗高变异幅度。在两种水分条件下,各树种的苗高地径变化趋势一致,均为700 mL水分条件下的值大于400 mL水分条件下的值。在两种水分条件下,生长状态最好的为楸树,梓树相对较差。
4.2 干旱胁迫对叶片水势的影响
在400 mL 水分条件下,除黄金树外,其他各树种的叶片水势(ΨL)都小于700 mL 水分条件下的值(P<0.01),但不同种减小的幅度不同(见表1)。从700 mL 到400mL 水分处理,黄金树的ΨL增加了24.77%,梓树、滇楸、灰楸和楸树的ΨL分别降低了16.32%、33.72%、15.85%和66.53%,表明不同种对干旱胁迫的适应能力有较大的差异。
4.3 干旱胁迫对气孔导度的影响
气孔导度(Gs)的日变化表现为单峰曲线,不同种达到最大值的时间各不相同(见表2)。在700 mL 水分条件下,黄金树、梓树和灰楸的Gs均在10:30 左右达到最大值;滇楸和楸树的最大Gs则出现在08:30 左右。在700 mL 水分条件下,灰楸的Gs峰值出现时间从10:30 左右前移到08:30左右,其他树种不变。
在干旱胁迫下各个种的最大Gs除黄金树外均表现出下降的趋势(见表1),然而不同树种下降的幅度差异较大。从700 mL 水分处理到400 mL 水分处理,黄金树的最大Gs增加了66.2%,梓树、灰楸、滇楸和楸树的最大Gs分别降低了45.0%、31.4%、20.0%和14.3%。同一水分处理下不同树种之间的最大Gs差异也较大。在700 mL 和400 mL 水分条件下,最大Gs的极差分别为:100.72 和200.79。由此表明在干旱胁迫下,各树种的最大Gs对胁迫的响应幅度也增大。
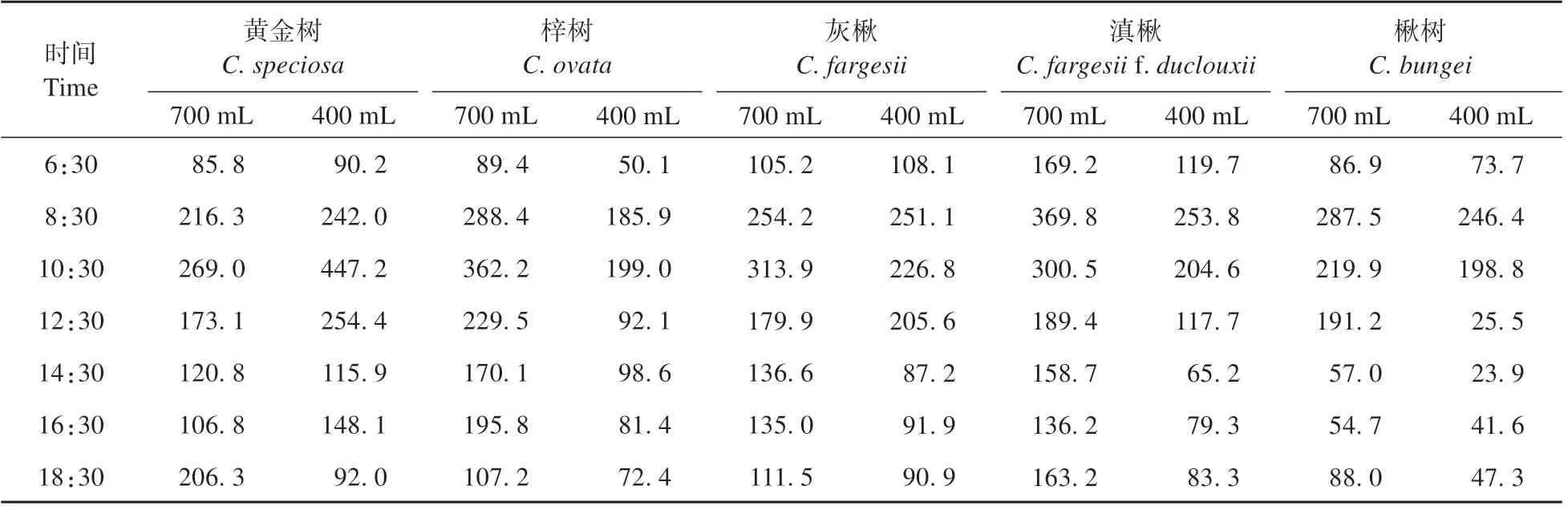
表2 干旱胁迫下不同种气孔导度日变化趋势Table 2 Diurnal variation trend of stomatal conductance of different species under drought stress
4.4 干旱胁迫对叶绿素相对含量和叶绿素荧光参数的影响
同一水分处理下不同种之间的叶绿素相对含量(SPAD)差异达到极显著水平(P<0.01)(见图2)。在两种水分条件下皆为滇楸和灰楸叶片的SPAD 值最高,楸树次之,而梓树和黄金树的SPAD值则相对较低。不同种的SPAD 值均随着干旱胁迫的加剧而升高。在700 mL 和400 mL 水分条件下,SPAD 值的极差分别为:13.27 和15.67。因此,随着干旱胁迫的加剧,各树种的SPAD 值随胁迫的变异幅度也随之增加。
除滇楸和楸树外,400 mL水分条件下,各树种的光系统Ⅱ(PSⅡ)最大光能转换效率(Fv/Fm)均大于700 mL水分条件下的值(见表3)。
4.5 不同水分梯度下比叶重的变化
比叶重(LMA)是指单位叶面积叶片的干重,是衡量叶片光合作用性能的重要参数[53]。不同种在相同的水分条件下其比叶重差异极显著(P<0.01),2 种水分条件下都是灰楸的LMA 显著高于其他4 个种,而其他4 个种之间则差异较小(见表3)。在2 种水分梯度下,除灰楸外,其他树种的LMA 都随胁迫加剧而升高,但变化均不显著。不同树种升高幅度具有差异,从700 mL 水分处理到400 mL水分处理,黄金树、梓树、滇楸和楸树的MA分别增加了0.5%、11.4%、7.6%和6.7%。

表3 各个种在2种水分条件下比叶重和最大光能转化效率的变化Table 3 Changes of specific leaf weight and maximum light energy conversion efficiency of each species under 2 waterconditions
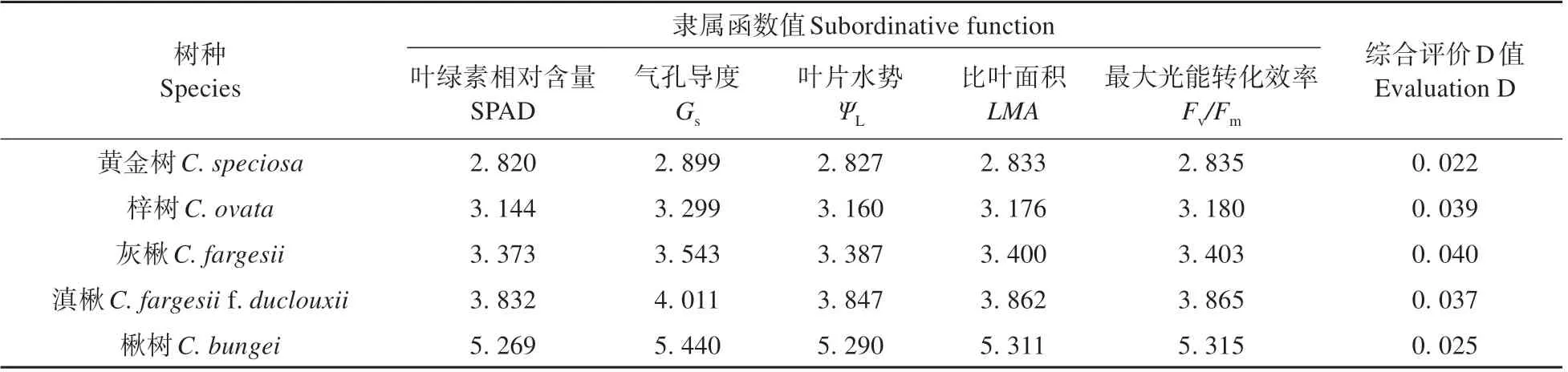
表4 干旱胁迫下各树种隶属函数值和综合评价D值Table 4 Value of subordinative function,evaluation index D of each species under drought stress
4.6 梓树属5个树种耐干旱评价
根据叶绿素含量、气孔导度、叶水势、比叶重和最大光能转换效率5个指标的相关指数,用隶属函数法对400 mL 水分条件下梓属5 个树种进行耐干旱评价(见表4)。5 个树种的综合评价D 值为0.022、0.039、0.040、0.037 和0.025。结果表明,其耐旱性由强到弱依次为灰楸>梓树>滇楸>楸树>黄金树。
5 讨论
在干旱胁迫下,植物会产生一系列生理上的变化,通过改变自身生理代谢功能来适应逆境[16]。植物叶片是与外界进行水分和气体交换,承担光合作用和蒸腾作用的主要场所,是植物对干旱胁迫较为敏感的器官[24],可以较好的反应植物对干旱胁迫的适应能力[23]。在土壤—植物—大气体系中包含了两个植物输水结构体系,包括:连接土壤与叶片的水分传输系统,和连接叶片与大气的叶片气孔[24]。在连接土壤与叶片的水分传输系统中,ΨL是植物水分亏缺的重要指标,高水势是维持正常水分传输的基本条件。随着干旱胁迫的加剧,土壤水势随之下降,因此植物通过迅速降低ΨL,来调整土壤与幼苗的水势,以此增加幼苗对水分的吸收能力[25]。干旱胁迫显著降低了‘波叶金桂’、山杏、香樟、杜英和刺槐等的ΨL[18,26~27]。在本研究中,随着干旱胁迫的加剧,除了黄金树外,各树种的ΨL也都表现出减小的趋势。这表明干旱胁迫中,除黄金树外,以上4 种梓属幼苗均可以通过降低ΨL从而吸收更多的水分,增加其抗旱性。除了增强对土壤中水分的吸收,植物叶片还会通过减少自身的蒸腾来维持体内的水分,即连接叶片与大气的输水结构体系。研究表明,Gs与ΨL存在相关性[18]。随着ΨL的降低,通过保卫细胞膨压的增大来控制气孔的开闭,以此降低叶片蒸腾作用,保持植物体内的水分,缓解干旱胁迫对幼苗的危害[29]。在本试验中,在干旱胁迫下,除了黄金树外,各树种的最大Gs均表现出下降的趋势,且浇水量的减少增加了各树种的最大Gs对胁迫响应的差异,这与张栋,张军等人的研究结果一致[28,30]。因此在干旱胁迫下,植物会逐渐关闭其气孔以减少水分的散失[31],干旱胁迫越严重,最大Gs越低。随着干旱胁迫的加剧,灰楸的Gs峰值出现时间从10:30 提前到8:30,表明灰楸通过调控气孔的开放,提前Gs峰值来适应干旱的环境,这与韩磊等人的研究结果一致[32]。
光合作用是植物获取能量的主要方式[33],植物可以通过调整自身的光合能力来适应环境的变化[34]。而叶绿素是光合过程中最重要的光合色素,直接关系着光合同化过程[35]。在以往的研究中,干旱胁迫会抑制叶绿素的合成并加快叶绿素的分解[36~37],导致叶绿素含量下降[38]。但也有不同结论,如PEG 诱导干旱胁迫提高了加勒比松(Pinus caribaea)的叶绿素含量,轻度干旱条件下柠条(Caragana microphylla)和紫穗槐(Amorpha fruticosa)的叶绿素含量有所增加[39]。而SPAD 值与叶绿素含量具有显著的相关性,在一定条件下具有较强的一致性[40~41]。在本研究中,随着干旱程度的加剧,各树种的SPAD 值均随之增加。随着干旱胁迫的加剧,各树种均通过增加叶片叶绿素含量来提高光合作用以此维持正常的生理功能。这与谭春燕等人在干旱胁迫下,植物需要保持较高的光合能力来维持生命活动这一结论相一致[16]。Fv/Fm为最大光化学效率,是表征植物是否受到光胁迫的一个有效指标[43~44]。何炎红,杨飞洋等人认为[42,44],干旱胁迫会降低PSⅡ反应中心的光能转化效率,降低Fv/Fm值,赵丽英等人则认为[45],Fv/Fm降低是由于植物可以做出一定的适应性调节,叶片内没有发生光抑制,而Fv/Fm的增加则为胁迫使植物体内发生光抑制,光合结构受到了一定破坏。在本研究中,梓属各树种的Fv/Fm受干旱胁迫影响均不显著。这可能是由于本试验的干旱胁迫程度较低,因此对各树种Fv/Fm的影响未达到显著程度。
LMA 与植物叶片的生理活动强弱相关,可以在一定程度上反映植物的抗旱性[14]。它把单位干重水平和单位叶面积水平的叶片指标联系在一起[46]。Witkowski、刘金环、李善家[47~49]等人研究认为,植物的LMA 越大,叶片越厚,其储水能力就越强,因此抗旱能力越强。这可以解释为,随着干旱胁迫的加剧,植物通过积累可溶性糖等渗透调节物质以降低胁迫对植物的伤害,增加叶片SPAD 值来提高光合作用,最终维持正常的生理功能,由此形成单位叶面积叶片厚度较大的叶片,即LMA 较大的叶片。在本研究中,除灰楸外,其他树种的LMA 都随胁迫加剧而升高,但差异不显著,说明其他各种均可通过渗透调节来降低干旱胁迫带来的伤害。
利用隶属函数法综合评价的结果为灰楸>梓树>滇楸>楸树>黄金树。各树种的苗高和地径均在700 mL 水分条件下达到最大值,说明5 个树种的最适水分条件均为700 mL。黄金树的ΨL和Gs均随着浇水量的减小而增加,700 mL 为黄金树正常生长的水分阈值,低于700 mL 水分条件时,黄金树会失去其水分与气孔调节能力,因此其抗旱能力较差。从苗高、地径的数值上,ΨL、Gs和Fv/Fm的变化上看,楸树为5 个树种中较耐旱树种;从Gs和SPAD值的变化,综合隶属函数分析,灰楸也为5个树种中较耐旱树种,这与Ma 等人的结论一致[50]。综上所述,梓属不同树种间有着不同的抗旱能力。综合分析,楸树和灰楸的抗旱能力较强,滇楸和梓树的抗旱能力次之,黄金树的抗旱能力最差。因此,渭北高原、关中平原、甘肃南部等地区可引种栽培楸树和灰楸,其有益于水土保持,具有较高的观赏、生态以及经济效益。
