东北地区李属(Prunes L.)植物导管分子形态结构研究
2021-02-26邹子瑜谷利伟张大维
邹子瑜 谷利伟 张大维*
(1. 哈尔滨师范大学,哈尔滨 150025;2. 黑龙江八一农垦大学,大庆 163319)
李属(Prunes L.)隶属于蔷薇科(Rosaceae)李亚科(Prunoideae)。该属植物全球约有150 余种,分布在除澳洲外各地,我国有50余种,全国均有分布;其中东北地区产10种,栽培15种[1]。李属植物具有较高的经济价值,桃(P.persica L.)、杏(P.vul⁃garis Lam.)、樱 桃(P.pseudocerasus(Lindl.)G.Don)等都是生活中常见的水果。稠李(P.racemosa(Lam.)Gilib.)、榆叶梅(P.triloba(Lindl.)Ricker)、樱花(P.yedoensis(Matsum.)Yu et Li)等具有较高的观赏价值。李属同时具有一定的学术价值:近年对李亚科各属的分类标准一直存在分歧:一种是“大属”分类法,1926 年由德国科学家A.Rehder提出,将核果类植物合并成李属,并将李属分为五个亚属:李亚属、扁桃亚属、樱亚属、稠李亚属与桂樱亚属,这种分类方法国内外采用者很多[2],本文也采用这种分类方法。另一种是由我国学者俞德浚提出的,将李亚科分为9 个小属。其中桃属(Amygdalus L.)最为进化,稠李属(Padus Mill.)与桂樱属(Laurocerasus Tourn. ex Duh.)相对最原始[3]。早期对李属植物的形态学研究以木材解剖学为主:认为李亚科和苹果亚科是蔷薇科中较为进化的亚科,且李亚科独立分支较早。并与苹果亚科亲缘较远[4~7]。随着近年来分子生物学的发展,对李属植物的分类研究也深入到了分子层面。Ma R.C.在对梨属与李属S-RNases cDNA 序列的研究中发现,扁桃仁(P.communis L.)与甜樱桃(P.avi⁃um(L.)Moench.)的S-RNases 基因没有特异性差异。这也可以成为李亚科大属分类的分子生物学佐证[8]。由于李亚科分类学仍存在分歧,而导管分子既存在一定微观分类学意义,又是反应生境状态的良好指标。因此,对蔷薇科植物导管分子的研究成为近年来的研究热点。我国学者尹秀玲曾对蔷薇科9 属12 种代表植物的导管分子进行观察,并找出了蔷薇科导管分子共性特征与属间差异[9]。郭学民等人发现李属砧木茎段到接穗茎段出现了导管宽度不匹配,说明即使为同属植物,不同种植物的导管直径在同生境下也存在着差异[10]。胡蓉在比较日本晚樱与樱桃导管分子的研究中发现,日本晚樱较樱桃更为进化,但他们个各性状在系统演化中不同步[11]。近年来对其他科属植物导管分子的研究以导管分子生理结构和不同植物导管分子形态结构差异为主。高建平、王彦涵等通过导管分子形态特征研究与年降水量比较,得出其与导管分子长度与直径的比值相关[12]。张大维等在研究桦木科(Betulaceae)与槭树科(Ac⁃eraceae)槭属(Acer Linn.)植物导管分子的过程中发现同科同属植物在结构形态上有一定共性,并发现导管分子的形态特征可能与环境因素有关[13~14]。
然而现阶段国内外对李属植物导管的分子的研究基本都停留于种间结构对比,并且同种植物没有在不同生境下进行结构与形态的分析。因此,本文选取了东北地区10 种当地自然生长的李属植物,并在不同生境多点采样。利用扫描电子显微镜进行观察,在研究导管分子结构的同时分析了生境因素对导管的影响,提供了微形态学证据。
1 材料和方法
1.1 材料
实验材料均采自中国东北地区自然环境生长的李属植物枝条(见表1)。选取一年生枝条的节间。每种植物分别采集自3个不同地理位置的生境下。
1.2 实验方法
采用植物导管树脂铸型法对导管分子内壁形态结构特征进行观察和测量。1 年生枝条自然干燥后,将周皮处理干净,切成10 mm×5 mm大小,浸泡于苯乙烯中放于真空抽气泵中抽出存留在输导组织中的空气,再浸泡4~6 周。渗透过苯乙烯的实验材料于70℃温箱中进行高温聚合,直到完全凝固为止。取出凝固好的材料,浸泡在以下2种溶液中使有机材料分解:①30%的过氧化氢和99.5%的冰醋酸1∶1的混合液。在60℃下浸泡1~2 h。②浸泡在98%的浓硫酸中10 min。更换溶液时需用蒸馏水冲洗后再浸泡在另一溶液中。两步骤轮流操作3~5 次,直至复型材料充分分散。暴露出的复型材料在70%的酒精中保存以待使用。样品材料经过完全处理后,稀释在70%的酒精中。
1.3 观察与测量
在解剖镜下用牙签、解剖针等较尖的用具将处理好的导管离析材料粘贴于扫描电镜的样品台上。再置于离子溅射仪中进行喷金导电处理,并在HITACHI S-4800 型扫描电子显微镜下(加速电压20 kV)观察。随机在扫描电镜的样品台上选择30个导管分子管长、尾长、宽度及端壁长度进行测量。根据端壁长度与导管宽度计算倾斜角正弦值,根据正弦值换算成角度(即角度数值越大面积越小)。

表1 实验材料信息Table 1 Experimental material information
1.4 数据分析方法
本文利用spss 软件对每组数据进行差异性分析与标准差分析。
2 观察结果
不同生境下的李属植物导管分子的管腔长度、宽度及端壁斜度角有较大的差别(见图1)。端壁倾斜角为端壁与直径的夹角度数。角度数值越小,说明端壁越倾斜,端壁面积越大。李属植物导管分子的类型有网纹、孔纹及螺纹3种形态;纹孔排列方式出现2 种分别是:对列、互列同时存在和仅存在互列式。李属植物导管分子均存在由管分子次生壁内侧具有螺旋状隆起的螺纹加厚(见表2)。由于同种植物不同生境下导管类型与结构特征并没有变化,故不区分采集地,仅以种名分类列表。
2.1 导管类型与穿孔板类型
研究中李属植物导管分子的导管形态类型共有3 种,网纹导管与孔纹导管普遍存在。仅山杏、欧李、东北李3种李属植物的导管分子中没有发现螺纹导管(见图版Ⅰ:1)。李属植物中普遍存在较为进化的孔纹导管。单穿孔板在该属孔纹导管中普遍存在,仅在樱亚属的黑樱桃、毛樱桃与郁李中发现梯状穿孔板。
2.2 纹孔的排列方式
李属植物导管分子纹孔排列方式为互列式的情况在所观察的植物中普遍存在,其中山杏、稠李、东北李以及毛樱桃为对列式、互列式同时存在(见图版Ⅰ:5~7)。其余6 种仅存在对列式(见图版Ⅰ:8~9)。
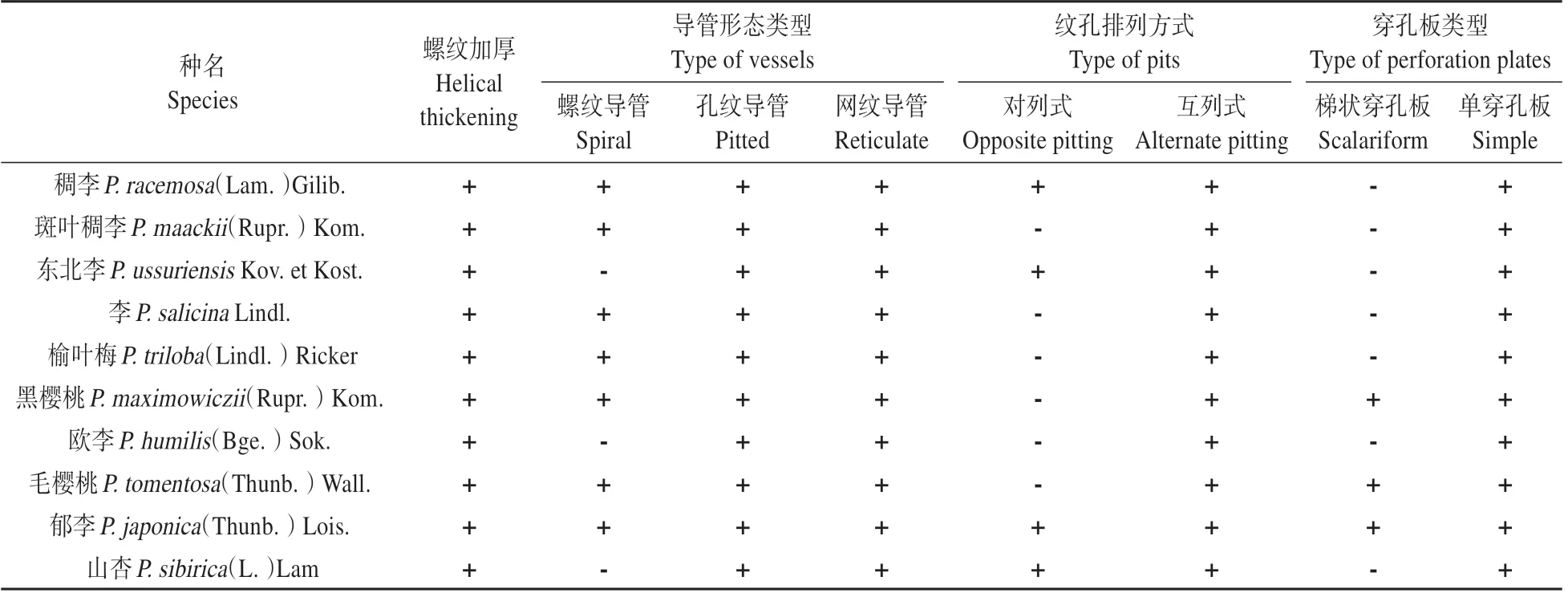
表2 东北地区植物导管分子形态特征Table 2 morphological characteristics of vessel elements from Northeast of China
2.3 导管管壁的螺纹加厚
所观察的李属植物导管分子的内表面均呈螺纹加厚(见图版Ⅰ:2)。螺纹型加厚会增加导管内壁的表面积,这可以提高导管对水分的附着力,从而提高水分运输效率,但不同生境下李属植物均存在螺纹加厚(见图版Ⅰ:4),且尾端也具有螺纹加厚与纹孔对(见图版Ⅰ:3)。
2.4 李属植物导管分子长度、端壁斜度角、宽度及尾端长度
所观察的10 种李属植物分别选择3 个不同的地区取样,共计30组样本。从数据分析可知,导管长度、端壁倾斜角与海拔和纬度正相关。并且在对榆叶梅、欧李、郁李跟山杏4 种植物的观察中发现,生长在长白山1 500 m 左右海拔的植物,纬度低于黑龙江地区,但导管长度与端壁角度与黑龙江地区的植物无显著差异,甚至高于黑龙江地区。导管宽度与海拔纬度呈负相关,即海拔、纬度越高导管宽度(即直径)越小。在对东北李的观察中发现,在海拔相对一致,纬度相对靠近的情况下,管长、宽度与端壁倾斜角度均无显著差异。在所观察不同生境下的李属植物中,尾端长度在种内相对稳定,无显著差异,但种间仍存在差异。
3 讨论
植物导管分子形态结构观察结果表明,同种植物在不同生境下形态(长度、宽度、端壁面积)上存在差异。但管壁螺纹加厚、尾端长度、导管类型,穿孔板类型以及纹孔式并没有出现变化。其中孔纹、网纹导管、螺纹加厚、互列纹孔式与单穿孔板是李属植物普遍存在的结构特征。梯状穿孔板仅在樱亚属植物中被观察到。但相对原始的螺纹导管存在在3种不同亚属的植物中。因此,我们认为导管分子的各结构的进化程度有所不同。对尾端长度的观察发现,同种植物尾端在不同生境下长度无变化,这说明尾端长度仅与进化程度相关。螺纹加厚在李属中普遍存在。内壁螺纹加厚是一种次生壁内侧产生的规则性凸出,它可以增强管壁对水的附着力,从而提高植物对水分的利用效率[15~16]。由于所观察的李属植物无论生境如何均存在螺纹加厚,所以推测这一形态应该是李属植物长期适应旱生环境而演化出来的,但这仍需要进一步研究。在对长度、宽度与端壁面积的测量中发现,李属植物在不同生境下导管分子会对环境产生一定的适应性变化。纬度与海拔越高,长度越长、宽度越小、端壁面积越小。这种变化是个体植物对环境适应的反馈,并不是适应环境而演化的结果。因此,我们认为李属植物导管分子的微形态结构具有十分重要的意义,既可以研究李属的系统学地位,又可以为李亚科的分类方法提供佐证。同时在对导管分子的环境适应性方面提供重要的参考依据。
导管的出现,是植物为适应陆地生境的一个重要标志。目前,国内外学者发现导管分子的形态结构具有较重要的系统发育学地位,导管分子中穿孔板的形态特征、导管类型及尾端长度是显示类群演化地位的重要微观指标[17~18]。梯状穿孔板作为较原始的穿孔板类型,常出现在较为原始的植物类群中,如毛茛科(Ranunculaeeae)等[19]。但在蔷薇科中较为进化的李属中仍有少数种存在梯状穿孔板(黑樱桃、毛樱桃与郁李)。且这些种都隶属于李属的樱亚属,但樱亚属的植物也存在较为进化的单穿孔板。因此,我们认为樱亚属的导管是东北地区李属中相对进化,但最为原始。由于导管分子管壁发育的先后和导管分子次生壁增厚的位置、方式不同,从而形成了不同类型的导管。螺纹、环纹导管常出现在植物发育的早期,而网纹与孔纹导管则出现在较为成熟的植物发育时期。并且国内外研究也表明导管类型也是分类学与系统发育学的重要指征[5,9,19]:环纹导管与螺纹导管较原始;网纹、孔纹导管则相对进化;梯纹导管则是较为原始类型的导管分子向孔纹导管进化的过渡类型。并且在所观察的植物中发现:不同生境下的同种植物导管类型相同,这与前人的研究相符合[20]。所观察的李属植物均存在孔纹导管与网纹导管;绝大多数李属植物均存在螺纹导管(仅东北李、欧李与山杏中没有)。这说明李属植物的导管类型以最进化的孔纹导管为主。多数种仍保留着较为原始、输水功能较弱的螺纹导管。欧李中既不存在螺纹导管,又没有发现梯状穿孔板。因此,我们认为欧李在李属中相对进化。本研究测量的尾端长度也支持这一结果。从导管类型与穿孔板类型来看李亚属与杏亚属相对较为进化,这也符合前人的结论。但他们仍存在相对过渡类型的纹孔式,即对、互列同时存在的纹孔排列方式。孔纹导管的纹孔式演化方式是由对列式向互列式方向进行的[21]。李属植物孔纹导管的演化已经处于较为进化的阶段,但尚存少许互列式纹孔同时说明着仍处于较为初级的进化阶段。综合来看,樱亚属的郁李中观察到了所有的原始结构特征,这显示着郁李是李属最原始的种。因此,我们认为李属植物的导管分子处于相对进化但较为初级的阶段。即已经普遍存在单穿孔板,孔纹导管等进化的标志,但仍存留如螺纹导管、梯状穿孔板等相对原始的结构。并且在相对进化的桃亚属中仍存在螺纹导管。但由于东北地区本地产的桃亚属植物相对较少,无法进行亚属内的比较,有待进一步研究。在之前的研究中,对导管分子尾端的研究基本停留在长度的相对性上:即尾端相对较长的导管分子较为原始,尾端较短的相对进化[13~14,22]。本研究则对李属植物导管分子的尾端长度进行了测量,认为李属植物导管分子尾端长度在24~36 μm,属于相对短的尾端长度。并且发现在不同生境下同种植物尾端差异不显著,长度较为稳定。这说明尾端长度可能仅与物种相关,与生境无关。由于樱亚属在李属中相对原始,但观察结果中樱亚属的毛樱桃尾端最长、同亚属的欧李、郁李却最短。推测樱亚属植物的导管分子不同特征可能在进化程度上不一致。但由于李属植物导管分子尾端长度整体相对稳定,因此本研究也支持将尾端长度作为衡量植物系统学地位佐证。但这也需要进一步测量蔷薇科其他各属植物后再进行比较分析。并且我们在观察中发现,导管分子尾端也有功能性结构(螺纹加厚与纹孔对),这也推测李属植物的尾端并不是无功能的盲端结构,而是可以参与横向水分运输的功能性结构。在环境适应性方面,导管为植物各个器官运输水分与无机盐,而植物导管其输水能力和导管分子的形态结构是密切相关的。端壁斜度角度大小直接影响管腔端壁截面的面积,端壁截面面积越大,运输水分阻力越大,输水效率越低。导管分子越长,单位长度内端壁数量越少,在增加管腔压力(即提高根系吸水能力)的同时,减少了端壁对水的阻力。导管直径越大,输水量越大,但越容易出现栓塞与空穴化[23~24]。但环境对长度与宽度的影响则有很多理解:Steven J.发现由寒带到亚热带植物导管分子的纹孔发生率明显上升。V.E.Ben⁃kova 在对西伯利亚的蔷薇科植物的木材解剖研究中发现,生长在亚北极地带的植物拥有较高的导管频率与较细的导管分子。刘树焕等则认为导管分子较长,端壁面积较小是导管分子适应高海拔等干旱条件的结果[22,25~26]。从导管分子的长度、宽度和端壁斜度角度数据分析结果显示,李属植物在高纬度高海拔地区为了适应生境提高了导管长度(即减少了单位长度穿孔板数量),减小了管腔直径(即降低了栓塞的发生率),从而提高了输水能力(即提高了水分利用率)。生长在吉林与辽宁的东北李,由于海拔与纬度变化幅度不大,所以在导管长度、宽度等形态上并没有显著差异,这也推测生境的纬度与海拔变化相对大时,导管分子形态会出现变化。但对比生长在黑龙江与辽宁的李时发现:在纬度相对较高的地区,到纬度高于一定程度时小幅度的纬度增加导管分子结构也可能出现差异。东北地区除长白山外,海拔均低于700 m,属于地平地带。比较适合同种植物在不同纬度下的生境比较。由于长白山均海拔1 500 m 以上,属于亚高山带。所以生长在长白山亚高山带的榆叶梅、欧李、郁李和山杏与生境在更高纬度的同种植物相比较,其导管长度相同甚至更长,宽度几乎相同,端壁面积相同甚至更小。因此,我们认为在高纬度、高海拔的生境下,植物为了更适应生境,导管分子形态会根据生境做出反馈:为提高输水效率,增加了导管长度进而减少了穿孔板数量。为降低空穴化发生率,降低了导管宽度,从而减少了导管栓塞的发生率。但成熟导管分子的宽度似乎存在最小极限,以至于高海拔情况下宽度并没有更大幅度的变小,但这仍需要进一步研究。
