伪狂犬病病毒FJSW 的分离鉴定及gG 基因序列分析
2021-02-23傅星源兆丰华生物科技福州有限公司福州350014
傅星源 兆丰华生物科技(福州)有限公司 福州 350014
伪狂犬病(Pseudorabies,PR)是由伪狂犬病病毒(Pseudorabies virus,PRV)引起多种家畜和野生动物以发热、奇痒(猪除外)和脑脊髓炎为主要症状的急性传染病。该病毒属于疱疹病毒科疱疹病毒亚科,又称猪疱疹病毒Ⅰ型,对猪的危害最为严重[1]。 该病易与其他疫病混合感染,曾经在世界上广泛分布,给养猪业造成的损失仅次于猪瘟和口蹄疫[2-3]。1990 年之后,由于伪狂犬病病毒基因缺失疫苗的使用,该病得到了控制,发病率大大降低,某些地区伪狂犬病病毒净化取得一定成效[4-6]。 但自2011 年以来,猪伪狂犬病又有反弹趋势,我国多个省、市、地区的猪场均发生了该病, 造成了一定经济损失[6-7]。 研究表明,2012 年后流行的伪狂犬病毒株gB、gC、gD、gE 等重要糖蛋白的氨基酸序列发生了突变[8-10]。本试验通过对福建某猪场疑似伪狂犬病死猪采集组织病料,进行伪狂犬病病毒分离鉴定, 成功分离出伪狂犬病病毒并进行测序。 与Genbank 中的参考毒株进行比较, 对福建地区伪狂犬病病毒株的分子流行病学及毒株溯源进行了初步探讨。
1 材料和方法
1.1 病料样品及主要试验材料 病料样品采集自福建邵武某猪场临床症状疑似伪狂犬病发病仔猪;2×GC Buffer、Taq DNA 聚 合 酶 为TAKARA 公 司产品;dNTP 为上海生工产品; 琼脂糖为Sigma 公司产品。
1.2 引物设计与合成 检测引物参照文献[11]的引物序列进行合成,预期扩增片段长度为347 bp;参照Genbank 中登录的伪狂犬病病毒基因组序列 (登录号NC006151)设计一对特异性引物,用于gG 基因扩增,预期扩增片段长度为1 501 bp,引物由上海生工生物工程有限公司合成。
1.3 DNA 提取及PCR 扩增 基因组DNA 提取按照金瑞鸿捷公司taco DNA/RNA 共同提取试剂盒说明书进 行。 PCR 反 应 体 系 为20 uL:2×GC Buffer 10 uL,dNTP (10 mmol/uL) 3 uL,ddH2O 3.8 uL,LA Taq DNA 聚合酶 (5 mmol/uL)0.2 uL, 上下游引物(100 uM)各0.5 uL,DNA 模板2 uL。反应循环参数:94 ℃、5 min;94 ℃、1 min,59 ℃、1 min,72 ℃、2 min,35 个循环;72 ℃、10 min。 PCR 产物用1%琼脂糖凝胶电泳检测, 并将阳性产物送至铂尚生物福州测序部测序。
1.4 病毒分离鉴定 将PCR 鉴定为阳性的脑组织匀浆液-20 ℃反复冻融3 次,12 000 r/min 离心10 min, 吸取上清通过0.22 um 滤器过滤除菌,取1 mL 接种单层Vero 细胞,37 ℃吸附1 h 后弃去接种液, 加入含有2%胎牛血清的DMEM 维持液,37 ℃继续培养,并设正常对照组。 定时观察细胞病变,病变达到80%时收获病毒液,连续传代3 次后经PCR 进一步鉴定。
1.5 gG 基因PCR 扩增、 序列分析 利用gG 基因特异性引物PCR 扩增得到目的基因片段,并送至铂尚生物福州测序部测序。 利用生物学分析软件DNAStar7 将分离株与GenBank 中登录的参考毒株gG 基因的序列进行比对及同源性分析,分析分离株的序列特征。

表1 序列分析用参考毒株
Bratha、Becker 和Kaplan 为欧美代表病毒株;Ea和LA 为国内以往分离株;其余为国内2012 年后分离株。
2 结果与分析
2.1 病料样品检测 将疑似伪狂犬病患猪的脑组织匀浆,提取总DNA 作为模板,利用检测引物进行PCR 检测。结果表明,3 头患猪的脑组织样品有特异性条带产生,大小与阳性对照条带相同,与预期结果相符,表明送检样品为伪狂犬病病毒阳性(见图1)。
2.2 病毒分离培养 阳性脑组织液经匀浆、过滤除菌后接种Vero 细胞,18 h 后即可观察到细胞圆缩、聚集,24 h 后可以观察到网状CPE,48 h 后细胞出现脱落、空斑(见图2)。 利用检测引物对培养物进行PCR 鉴定,能够扩增到特异性目的片段,而利用猪瘟病毒、猪圆环病毒、猪繁殖与呼吸综合征病毒等特异性引物进行PCR 扩增均为阴性反应。表明从患猪脑组织中分离出的是伪狂犬病野毒株。
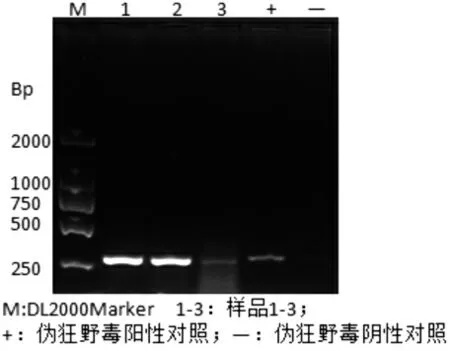
图1 患猪脑组织样品PCR 鉴定

图2 分离株在Vero 细胞上的病变
2.3 核苷酸序列同源性比较分析 对病变的Vero细胞营养液提取DNA, 用伪狂犬病病毒gG 基因特异性引物进行PCR 扩增,获得与预期大小一致的约1 500 bp 片段,并对其进行测序。结果表明,FJSW 株的gG 基因测序片段长度为1 501 bp。选择GenBank上参考毒株的gG 基因序列进行同源性对比,FJSW毒株与参考毒株核苷酸同源性为98.9%~100%。 其中与国内2012 年后分离毒株的同源性较高,为99.7%~100%,其中与JS-2012 株的同源性为100%;与欧美分离株的同源性最低,为98.9%~99.1%,其中与Kaplan 株的同源性为98.9%;与国内以往分离株的同源性居中,其中与LA 株的同源性为99.7%,与Ea 株同源性为99.9%。 与各毒株核苷酸的同源性比较见表2。

表2 核苷酸序列同源性比较分析
2.4 基因进化树分析 基于gG 基因核苷酸序列的遗传进化分析显示,FJSW 株与JS-2012 株处于独立的小分支上, 形成单独的1 个小亚群, 亲缘关系最近,该结果与同源性分析结果相符;与国内2012 年后 分 离 株HeN1、BJ/YT、TJ、XiangA 处 于 同 一 分 支中,表明FJSW 株属于伪狂犬病病毒变异毒株;而国内2012 年后分离株又与国内以往分离株LA、Ea 处于同一大分支上,形成了一个中国毒株进化群。欧洲分离株Kaplan、Bartha 与美洲分离株Becker 处于另一大分支上。
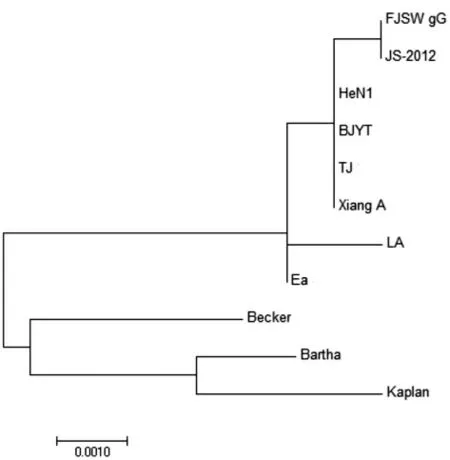
图3 基于gG 基因核苷酸序列的遗传进化分析
3 讨 论
自2012 年以来, 我国多地相继暴发猪伪狂犬病,造成了重大的经济损失[6-7]。 研究表明,伪狂犬病病毒变异毒株某些重要的糖蛋白在多个方面发生变异,存在毒力增强的情况[8-10]。 gG 糖蛋白位于PRV基因组US 区域,全长1 494 bp,可编码498 个氨基酸序列,为非结构蛋白,当病毒在细胞内合成该蛋白后被宿主细胞分泌到细胞外。 尚未明确gG 糖蛋白的毒力影响, 但gG 糖蛋白影响伪狂犬病病毒的吸附、扩散、穿透作用,在病毒与宿主细胞之间的扩散具有连接作用,可以抑制细胞迁移,对宿主细胞的凋亡有一定影响,在免疫逃避中扮演重要角色[12]。
为探明FJSW 株与参考株之间的亲缘关系,本试验对FJSW 株的gG 基因进行PCR 扩增、测序,与Genbank 上已发布的参考毒株进行同源性比对和多序列比对分析,并绘制遗传进化树。同源性比对结果表明,FJSW 株与2012 年后分离毒株在核苷酸水平上同源性较高,与国外经典毒株同源性较低。遗传进化树分析表明, 国内毒株均同属于同一个分支,且2012 年后分离毒株的遗传关系更近,FJSW 株与2012 年后分离毒株位于同一分支,与JS-2012 亲缘关系最接近; 国内毒株与欧美分离株的遗传关系较远。
综上所述,FJSW 株与2012 年后分离的变异株亲缘关系较近,属于PRV 新流行毒株。在gG 基因上2012 年后分离株与之前的国内外分离毒株存在不同程度的差异,但这些核苷酸水平上的差异对PRV变异株毒力以及免疫原性的影响有待进一步研究。
