LPS 诱导仔猪腓肠肌相关microRNA在不同时间点的mRNA 表达
2021-02-22黄兴法王秀英刘玉兰汪文俊
黄兴法 , 徐 鑫 , 张 晶 , 王秀英 , 刘玉兰 , 汪文俊 , 康 萍
(1.中南民族大学武陵山区特色资源植物种质保护与利用湖北省重点实验室,湖北武汉 430074;2.武汉轻工大学动物营养与饲料科学湖北省重点实验室,湖北武汉 430023)
随着养殖规模的扩大, 如何降低外界环境因素对养殖业带来的经济损失一直是研究者关注的重点。 环境中的微生物,如细菌、真菌和病毒等均会导致仔猪在饲养过程中产生免疫应激反应,进而诱导动物炎症。 脂多糖(LPS)是革兰氏阴性菌外膜的主要成分,在组织细胞中,能快速诱导促炎细胞因子的产生,进而引起炎症反应(Irtegun 等,2016)。miRNA 是一种约含22 个核苷酸的内源性非编码单链小RNA 分子,在大多数真核生物中存在高度保守性 (Geng 等,2014)。 许多哺乳动物miRNAs 可以调节免疫反应和炎症反应 (Qiu 等,2019;O"connell 等,2012)。 腓肠肌是小腿后面浅层的大块肌肉,其组织中的蛋白酶体标志物、氧化应激、炎症、线粒体呼吸链(MRC)活性、耗氧量等指标一直被广泛研究(Barreiro 等,2016)。 虽然目前对LPS 刺激后诱导的机体组织编码基因关键信号通路研究很多, 但对相关非编码基因在炎症反应中调控的研究并不多。因此,本试验旨在研究仔猪腓肠肌相关miRNA 在发生炎症断奶仔猪的基因表达情况,为后续研究miRNA 在肌肉组织中的炎症调控作用提供科学依据。
1 材料与方法
1.1 试验设计 选择体重为(7.1±0.9)kg 的杜×长×大断奶仔猪42 头,饲喂基础饲粮14 d 后,设置对照组(注射 LPS 之前 0 h)和 LPS 组(注射 LPS 之后 1、2、4、8、12、24 h)。分别按体重 100 μg/kg 腹腔注射生理盐水和LPS,然后屠宰取腓肠肌样品待测。
1.2 试验饲粮 饲粮配方采用玉米-豆粕型基础饲粮,参照NRC(1998)仔猪营养需要进行配制,具体组成及营养水平见表1。
1.3 试验材料 本试验注射LPS 的血清型、来源、配制方法与汪洋等(2019)的一致。
表1 基础饲粮组成及营养水平(风干基础)
1.4 饲养管理 动物饲养试验在动物营养与饲料科学湖北省重点实验室进行。 本试验饲养管理方法参照陈会甫等(2018)的方法进行。
1.5 测定指标与方法 用 RNAiso Plus 试剂(#9108,TaKaRa) 提取腓肠肌组织总 RNA。 根据PrimeScriptRT Reagent Kit with gDNA Eraser(#RR047A,TaKa-Ra)说明书进行基因组 DNA 去除,并用mirVanaTMqRT-PCR 引物套装(吉玛基因,上海)和 SYBR Premix Ex TaqTM(Tli RNaseH Plus)qPCR Kit (#RR420A,TaKaRa) 进 行 成 熟miRNA 定量检测。 以 U6 为内参,并参照 Livak 等(2001)的 2-△△CT法进行计算。
1.6 统计学分析 试验采用SPSS 22.0 软件进行单因素方差分析和Duncan’s 多重比较,结果采用“平均值±标准误”表示,P <0.05 表示差异显著。
2 试验结果
LPS 刺激对断奶仔猪腓肠肌miRNA 相关基因表达量的影响结果见表2。 与对照组相比,LPS刺激后2 h,仔猪腓肠肌miRNA-10b 的mRNA 表达量显著降低 (P < 0.01);LPS 刺激后 1 h, miRNA-1 和miRNA-206 的mRNA 表达量显著降低(P < 0.01);但 LPS 刺激后 miRNA-133a 的 mRNA表达量无显著变化(P > 0.05)。
3 讨论
给断奶仔猪注射一定剂量的LPS 是建立仔猪炎症的良好模型(Zhu 等,2013)。研究表明,LPS刺激后4 h 可诱导肌肉炎性细胞因子大量表达(李先根等,2019)。 还有研究报道,先天性免疫细胞可以处于不同的状态, 具有不同程度的促炎和抗炎表型(Hanke 等,2013;Stephanie 等,2009),并伴随着对宿主防御和炎症的不同后果(Marrotte等,2010)。 汪洋等(2019)的研究也表明,LPS 刺激激活了炎症信号通路, 使炎性细胞表达量显著上调,并在不同时间点,这些基因呈现先上升后逐渐下降的变化。 随着现在对miRNA 的深入关注,发现不同的miRNA 对机体组织炎症有着不同的调控功能。 例如:通过有效下调miRNA-DAB2IP 可以增强癌细胞的运动和侵袭性,促进NF-κB 信号的激活, 进而促进促炎症和促血管生成因子的表达(Bellazzo 等,2018)。 miRNA-19b 通过抑制胸腺基质淋巴生成素下调小鼠哮喘模型的STAT3 信号来减少氧化应激程度(Ling 等,2017)。 miRNA-125b 上调可激活NF-κB 途径促进类风湿关节炎(RA)炎症(Zhang 等,2017)。 此外,炎症相关的miRNA 在牛乳中的表达水平受到乳腺炎的影响,牛奶中的miRNA 有可能作为牛乳腺炎的生物标志物(Lai 等,2017)。 以上研究表明,机体内这些小分子RNA 虽然不能编码蛋白质,但是其也能够调控机体的炎症相关信号通路。 因此本试验的目的是研究LPS 处理前后不同时间点不同miRNA的表达,为后续研究miRNA 在肌肉组织中的炎症调控作用提供科学依据。
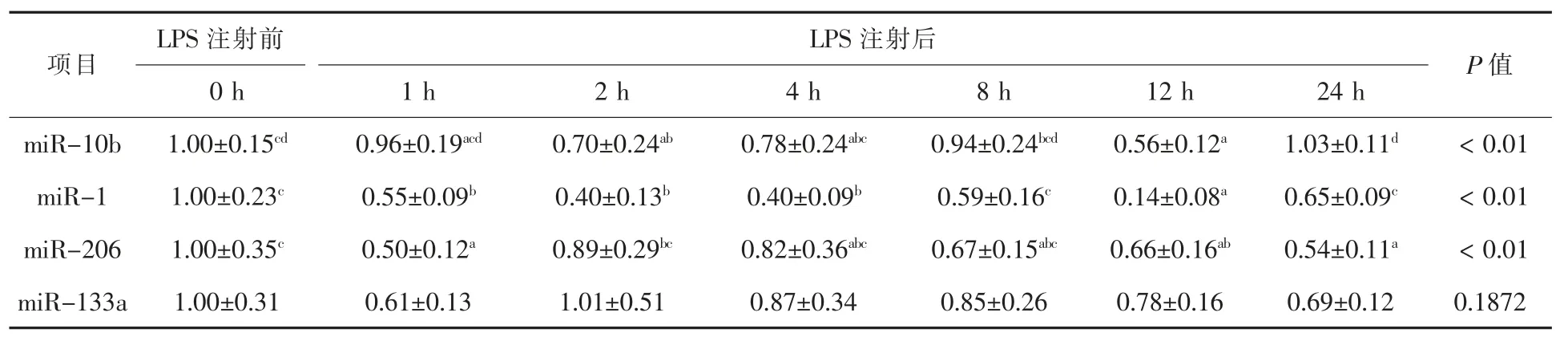
表2 LPS 处理前后不同时间点miRNA 表达
miRNA-10b 是最先被证明在癌症中有异常表达的 miRNA 之一(Mal 等,2017)。 迄今为止,已有超过100 项关于miRNA-10b 与18 种癌症转移的研究。 Sheedy 和 Medarova(2018)认为 miRNA-10b 作为癌症治疗和诊断靶点的潜力是非常重要的。此外,miRNA-10b 还与骨形成呈正相关,可促进体外成骨和抑制体外成脂(Li 等,2018)。 在本研究中,LPS 刺激 2 h 后,miRNA-10b 表达显著降低, 说明miRNA-10b 可能对动物早期免疫应激后起到了负调控的作用。miRNA-1 是最先被证明在肌肉细胞衍生物中特异表达的miRNA 之一(Nicholas 等,2005)。 截止到目前,已有 50 余项关于miRNA-1 在肌肉相关信号通路调控的研究。Cai 等(2015)认为,miRNA-1 与转录后调控多种与心肌电活动相关的离子通道和蛋白有关。此外,miRNA-1 还对心肌细胞肥大、细胞外基质沉积以及心脏重塑至关重要 (Luo 等,2018)。 在本研究中,LPS 刺激 1 h 后, 腓肠肌中 miRNA-1 表达显著降低, 说明miRNA-1 可能对动物早期免疫应激也起到了负调控的作用。miRNA-206 是最先被证明在小鼠骨骼肌中有异常表达的miRNA 之一(Mccarthy 等,2007)。迄今为止,已有 20 余项关于miRNA-206 在肌肉组织中的研究。 Zhang 等(2019) 认为,miR-206 在骨骼肌生长发育调控中非常重要。此外,miRNA-206 还与乳腺癌有关,可作为预测乳腺癌患者预后的重要指标 (Xing 等,2016)。 在本研究中,LPS 刺激 1 h 后, 腓肠肌中miRNA-206 表达显著降低, 说明 miRNA-206 可能对动物早期免疫应激起到了负调控的作用。miRNA-133a 首次被证明是在成人骨骼肌中有异常表达的 miRNA 之一(Mccarthy 等,2007)。 迄今为止, 已有200 余项关于miRNA-133a 在肌肉中差异表达的研究。Ramos 等 (2018)认为,miRNA-133a 通过耗尽肌肉细胞内储存并释放其对靶基因表达的抑制作用,潜在地介导运动的生理适应。此外,miRNA-133a 还与腰椎骨密度呈负相关,通过促进破骨细胞分化参与了绝经后骨质疏松症的调节(Zhong 等,2018)。 在本研究中,LPS 刺激 1 h后,miRNA-133a 无显著变化,说明在仔猪应激早期可能没有参与机体的炎症调控。 综上, 在本试验中, 通过建立仔猪LPS 炎症损伤模型显示,仔猪腓肠肌组织中的 miRNA-10b、miRNA-1、miRNA-206 的基因表达量均显著下调, 但miRNA-133a 的基因表达量无显著变化,这表明在LPS 刺激后同一个miRNA 在不同机体组织炎症反应中的表达也不一致。
4 结论
LPS 刺激下诱导腓肠肌相关基因miRNA-10b、miRNA-1 和 miRNA-206 在不同时间点均有显著降低,表明这些miRNA 可能参与了肌肉炎症早期相关信号通路的调控,可为后续miRNA 在肌肉组织中具体调控作用提供科学依据。
