臭氧处理对花生蛋白结构性能的影响
2021-02-06严永红郑召君李进伟
严永红 郑召君 林 叶 李进伟
(无锡中粮工程科技有限公司1,无锡 214035) (江南大学食品学院2,无锡 214122)
作为我国重要的油料作物,花生年产量已逾1700万t,其中约50%以上用于榨油[1]。而榨油后副产物——花生蛋白质含量高,约占40%~50%,其必需氨基酸含量约占总蛋白的11%左右,是一种优质的植物性蛋白资源[2-4]。然而花生蛋白在加工利用过程中易受氧化剂的影响,蛋白质结构特性发生改变,进而影响相关产品的品质。因此,解析氧化剂对花生蛋白结构特性的影响对改善产品加工品质具有重要意义。
作为一种可接触型食品氧化剂和消毒剂,臭氧自从1977年被美国FDA公认为食品类安全添加剂以来,被广泛应用于食品工业领域[5]。臭氧处理可以改变麦谷蛋白的二级结构,提高其蛋白分子尺寸及紧密度,进而改变面团的流变特性,并对小麦粉具有增白效果[6, 7]。臭氧具有杀菌消毒作用,安全无污染,也常被应用于水产品的加工与贮藏[8]。此外,有研究表明臭氧有助于氧化降解花生仁、玉米、开心果中的黄曲霉毒素[9, 10],且其对花生仁的降毒效果明显优于小麦粉[11]。林叶等[12]发现臭氧处理可降低花生中黄曲霉毒素B1(140 μg/kg),使其低于国家标准GB 13078—2001的限量要求,但臭氧处理会导致蛋白体外消化率与溶解性、乳化性、凝胶性等功能特性的降低。因此,臭氧因其氧化、消毒和降毒功能而被广泛应用于食品加工过程,但臭氧处理对花生蛋白结构特性的影响尚不明晰。
本研究以花生蛋白为研究对象,探讨臭氧处理前后蛋白的氨基酸组成、空间结构、巯基以及羰基变化,进而深入探讨臭氧处理对花生蛋白结构特性的影响,为臭氧应用于花生提供参考。
1 材料与方法
1.1 材料与试剂
花生、丙烯酰胺、1-苯氨基萘-8-磺酸、(ANS)、四甲基乙二胺(TEMED)、5,5′-二硫代二硝基苯甲酸(5,5′-dithiobis(2-nitrobenzoic acid),DTNB)、十二烷基硫酸钠等试剂均为分析纯。
1.2 仪器与设备
HY-003臭氧发生器,Jasco J-715圆二色谱仪,Waters高效液相色谱仪,F-7000荧光光谱仪,FE20 pH计,HH-4数显恒温水浴锅,TA Q100差示扫描量热仪。
1.3 试验方法
1.3.1 臭氧处理花生粕
花生粕样品(100 g)置于通入100 mg/L臭氧的空气洗瓶中,此时气体流量为4.0 L/min,每充气两分钟充分振摇一次,并于臭氧处理0、2、5、10、20 min时收集样品,即为不同臭氧处理时间下的花生粕样品。
1.3.2 提取花生蛋白
参考Reddy等[13]花生蛋白提取方法,并稍作修改。处理前后的花生样品溶于去离子水以制备浓度为10%的溶液,随后加入2 mol/L NaOH溶液调节至pH 9.0。室温震荡1 h后9 690 g、4 ℃离心30 min,收集上清液。而后加入2 mol/L HCl调节pH至4.5以沉淀蛋白,9 690 g、4 ℃离心20 min,收集沉淀后加水复溶,并调节溶液至pH 7.0。冷冻干燥获得花生蛋白样品,4 ℃储存备用,并采用凯氏定氮法测定蛋白含量。
1.3.3 氨基酸组成测定
150 mg花生蛋白样品与 8 mL 6 mol/L HCl 混合均匀,置于110 ℃条件下水解约22 h。水解液过滤、离心,收集上清液,采用高效液相色谱法测定样品中氨基酸含量。
1.3.4 巯基与二硫键含量测定
参照Beveridge等[14]报道的方法,测定臭氧处理前后花生蛋白的游离巯基(SH)与二硫键(-S-S-)含量。按照式(1)计算总巯基(Total SH)含量。
总巯基含量/(μmol/g Pro)=2×二硫键含量+游离巯基含量
(1)
1.3.5 羰基含量测定
参考Levine等[15]的方法并稍作修改。将1.0 mL花生蛋白溶液(2 mg/mL)与3.0 mL 10 mmol/L 2,4-二硝基苯肼溶液(含2 mol/L HCl)混合均匀,室温条件下避光反应1 h。在上述混合液中加入1 mL三氯乙酸(50%)后,10 000 g离心20 min收集沉淀,并用乙醇-乙酸乙酯溶液(1∶1)反复洗涤沉淀3次。之后加入3 mL盐酸胍溶液(6 mol/L),37 ℃水浴20 min溶解沉淀,并于367 nm波长处测定所得溶液的吸光值。
1.3.6 差示量热扫描
采用差示扫描量热仪(DSC)测定花生蛋白的热性质。2.5 mg样品加入10 μL磷酸盐缓冲液(0.01 mol/L,pH 7.0)混匀加入铝盘并密封,进行DSC扫描,温度扫描范围20~110 ℃,升温速率5 ℃/min。利用Universal Analysis 2000 软件处理图谱获得花生样品的变性温度(Td)和总变性焓(ΔH,J/g干物质)。
1.3.7 表面疏水性测定
参照Kato等[16]方法,利用ANS荧光探针法测定臭氧处理前后花生蛋白的表面疏水性。采用磷酸盐缓冲液(10 mmol/L,pH 7.0)配制一系列花生蛋白溶液,溶度分别为0.05、0.1、0.2、0.5、1 mg/mL。随后将20 μL ANS溶液加入至4.0 mL蛋白溶液中,混合均匀,利用荧光光谱仪进行荧光强度的测定,激发波长和发射波长分别为365、484 nm。蛋白质表面疏水性则表示为荧光强度随蛋白质浓度变化曲线的初始斜率。
1.3.8 圆二色(CD)光谱测定
参考Zhao等[17]方法并稍作调整。10 mmol/L磷酸盐缓冲液(pH 7.0)溶解花生蛋白制备浓度为0.25 mg/mL的样品溶液,并利用CD光谱仪分析其CD特性。主要参数为:扫描波长190~250 nm,扫描宽度1.0 nm,扫描速率100 nm/min,扫描频率5次。记录光谱并分析花生蛋白的二级结构。
1.3.9 十二烷基硫酸钠一聚丙烯酰胺凝胶电泳(SDS-PAGE)分析
参照Jiang等[18]方法并稍作修改,分别采用还原型和非还原型SDS-PAGE方法测定花生蛋白的分子量分布。分离胶和浓缩胶浓度分别为12%和4%,上样10 μL,在恒流模式下进行蛋白电泳。与非还原型SDS-PAGE相比,还原型SDS-PAGE蛋白电泳样品另外加入1 mmol/L β-巯基乙醇。电泳结束后,采用考马斯亮蓝染液对蛋白胶进行染色,随后利用甲醇-醋酸溶液脱色,并用凝胶成像仪采集电泳图像。
1.4 数据统计分析
本研究试验组内测定3次平行,结果采用“均值±标准误”形式表示,并采用Origin 8.0统计软件进行数据统计分析并作图,不同的小写字母表示P<0.05。
2 结果与讨论
2.1 臭氧处理对花生蛋白氨基酸组成的影响
蛋白质一级结构主要表现为氨基酸组成与含量,氨基酸组成的差异影响蛋白质的结构与功能[19]。众所周知,加工处理过程可能会造成氨基酸的差异,这与底物蛋白、氨基酸类型以及加工方式有关[20]。因此,臭氧处理对花生蛋白氨基酸组成的影响见表1。花生蛋白中占比最高的必需氨基酸为苯丙氨酸,其含量随臭氧处理时间延长而呈下降趋势,这可能是因为部分苯丙氨酸被臭氧氧化成醌类化合物[21]。类似的是,甲硫氨酸和半胱氨酸等含硫氨基酸含量也呈现出时间依赖性,随臭氧处理时间增加而逐渐下降。推测其原因,可能是因为臭氧氧化含硫氨基酸的巯基生成二硫化物类,从而导致甲硫氨酸和半胱氨酸分别转化为砜和胱氨酸[21, 22]。相比较而言,其他氨基酸含量变化不明显,这可能与氨基酸种类以及氨基酸残基所处蛋白质中的位置有关,氨基酸残基蜷缩于空间结构内部,呈现相对稳定状态而不易被臭氧氧化或对臭氧作用不敏感。

表1 臭氧处理前后花生蛋白的氨基酸组成/%
2.2 臭氧处理对花生蛋白巯基与二硫键的影响
臭氧处理导致花生蛋白甲硫氨酸和半胱氨酸含量下降,这可能与其内部的巯基和二硫键息息相关,而其含量的变化标志着蛋白质氧化[23]。由图1可知,随着臭氧处理时间延长,巯基含量显著性下降(P<0.05),而二硫键含量则呈现明显上升趋势。究其原因,可能是臭氧处理过程中形成氧自由基,氧化巯基而形成二硫键,导致巯基下降而二硫键升高的现象[24];此外,臭氧可以促进巯基与其他化合物发生反应,从而降低游离巯基含量[25]。臭氧处理后花生蛋白中二硫键含量的不断增加也许与巯基以及含硫基团被氧化有关,或者是两分子的半胱氨酸侧链上的巯基发生聚合形成胱氨酸,从而提高了二硫键含量。相似地是,Liu等[26, 27]研究发现,氧自由基导致氧化蛋白质的二硫键含量升高而巯基含量下降。值得注意的是,花生蛋白质中二硫键约是同等条件下巯基含量的几十倍,因此总巯基与二硫键含量变化趋势相似,随臭氧处理时间延长而逐渐增加。
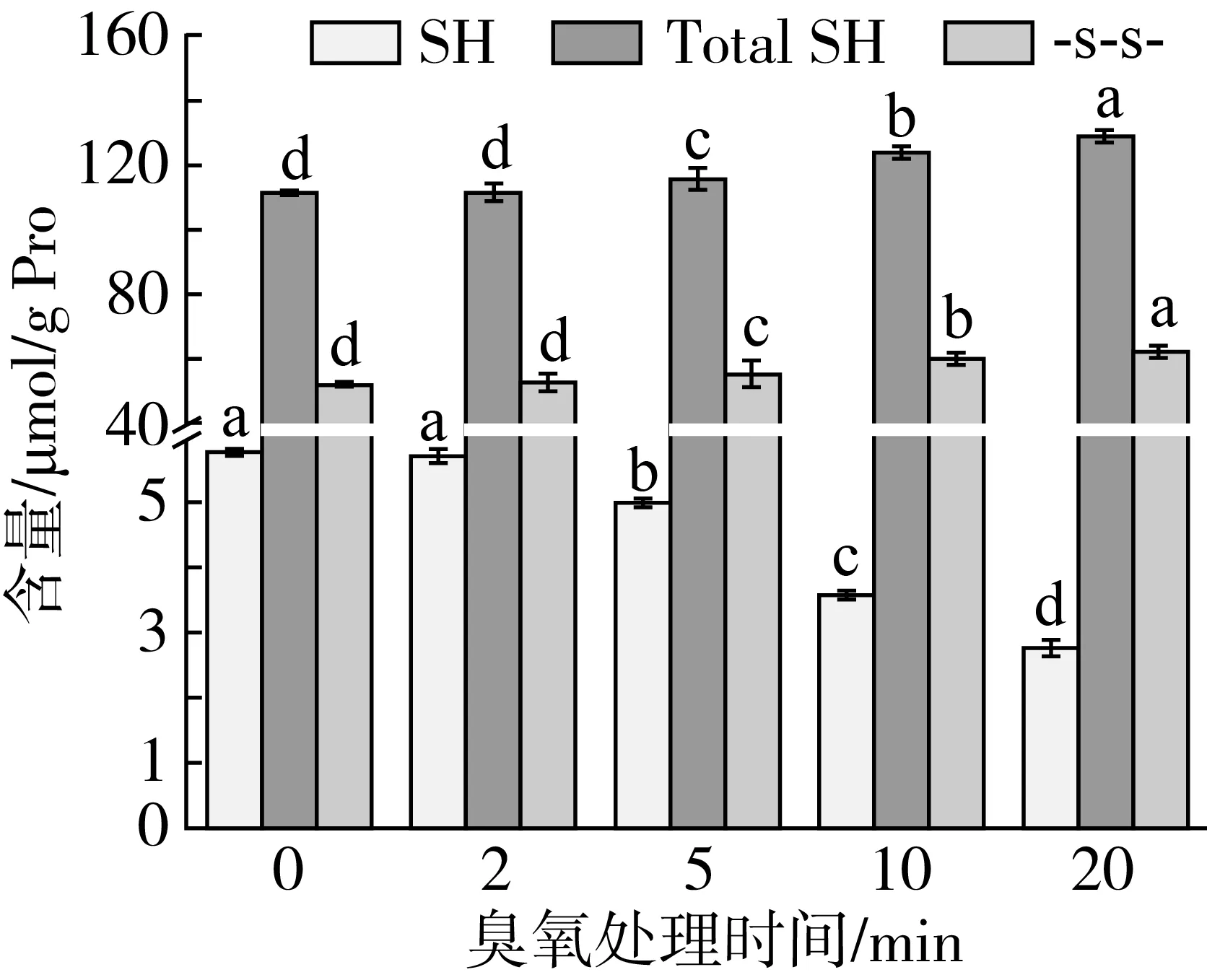
图1 臭氧处理对花生蛋白巯基和二硫键含量的影响
2.3 臭氧处理对花生蛋白羰基含量的影响
蛋白质中的氨基酸侧链基团易被氧化剂氧化成羰基衍生物,因此羰基含量的变化直接与蛋白质氧化程度相关联[28]。由图2可知,与对照组相比,臭氧处理导致花生蛋白质中羰基含量显著提升,且随臭氧处理时间延长而不断上升。这说明臭氧导致部分蛋白发生氧化反应,产生氧自由基,而活性氧则会攻击氨基酸残基中游离氨基或亚氨基生成羰基,或是高活性氧自由基导致肽链骨架断裂而生成羰基衍生物[29]。此外,样品中残留脂质在臭氧作用下氧化形成高活性的自由基,从而与花生蛋白残基反应,导致蛋白质中羰基含量增加[30]。
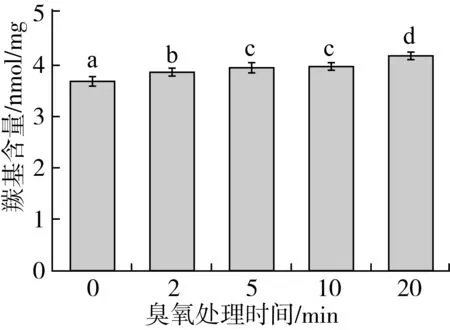
图2 臭氧处理对花生蛋白羰基含量的影响
2.4 臭氧处理对花生蛋白热性质的影响
臭氧处理可导致蛋白构象变化,从而改变了蛋白质的热稳定性。与对照组相比,臭氧处理导致花生蛋白的初变性温度(Tm)以及变性温度(Td)升高,进而引起吸热焓增加,表明臭氧可使蛋白质热稳定性提高,蛋白质结构舒展变性需要更高的能量(表2)。随着臭氧处理时间延长,花生蛋白的变性温度呈上升趋势,这可能是因为臭氧处理导致二硫键共价聚集形成更紧密的蛋白结构,进而强化了蛋白热稳定性[31]。相同地是,在0~10 min时,花生蛋白的吸热焓与臭氧处理时间呈正相关,但处理20 min后的花生蛋白吸热焓略有下降。原因可能是长时间臭氧处理破坏蛋白有序的折叠结构,这与臭氧作用于小麦蛋白的结果相似[32]。
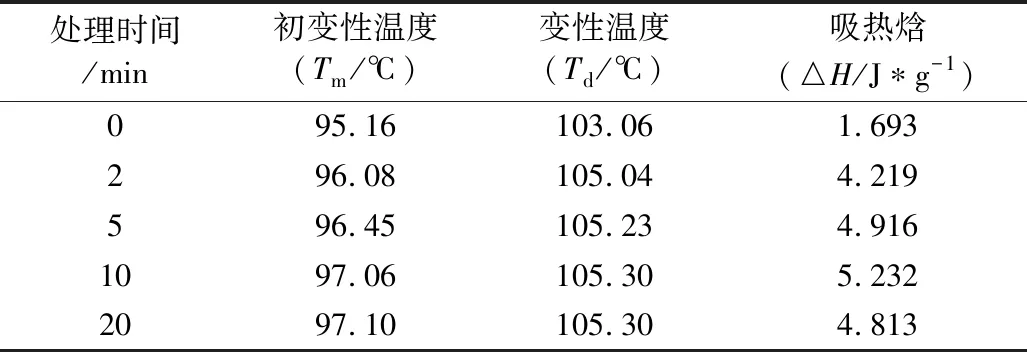
表2 臭氧处理前后花生蛋白的变性温度及变性焓
2.5 臭氧处理对花生蛋白表面疏水性的影响
如上所述,臭氧处理导致苯丙氨酸和蛋氨酸等疏水性氨基酸含量有所下降,由此推测臭氧可能影响花生蛋白的表面疏水性。由图3可知,0~10 min臭氧处理可导致花生蛋白的表面疏水性下降,处理时间与表面疏水性呈负相关线性关系(R2=0.988 1)。由此可推断,臭氧逐渐氧化花生蛋白表面的疏水性基团,降低疏水性;或者是臭氧作用加剧了花生蛋白内部的疏水性基团的过度暴露,基团之间彼此相互作用,导致部分疏水性位点被遮盖,从而降低了疏水性。而相反地是,臭氧处理20 min的花生蛋白表面疏水性反而显著高于处理10min的样品,表明过度臭氧处理加速暴露蛋白内部的疏水性基团;也可能是长时间臭氧氧化生成具有疏水特性的化合物,反而增加了花生蛋白的表面疏水性。
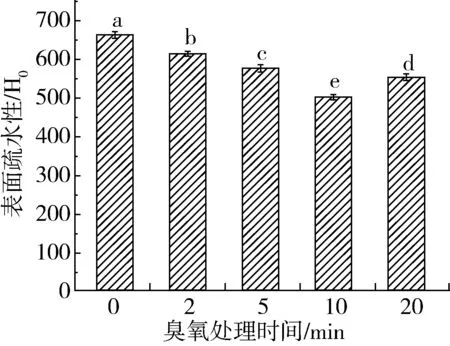
图3 臭氧处理对花生蛋白表面疏水性的影响
2.6 臭氧处理对花生蛋白二级结构的影响
CD光谱是表征蛋白质二级结构变化的一种精准、快速技术。如图4所示,对照组(臭氧处理0 min)在195~200 nm呈现正峰模式,210 nm左右则显示出负峰,表明花生蛋白具有明显的α-螺旋/β-折叠结构。而经过臭氧处理后,花生蛋白的CD光谱强度呈现随处理时间延长而逐渐降低趋势,正峰波长随处理时间延长而发生蓝移,负峰波长则发生红移现象。由此推测,臭氧可诱导花生蛋白二级结构的变化,有序的蛋白结构逐渐失稳转向不规则的二级结构。从表3也可以看出,臭氧处理导致花生蛋白中α-螺旋、β-折叠以及β-转角结构的比例明显减少,随处理时间延长而逐渐下降,相反地,无规则卷曲结构所占比例呈现显著上升趋势。这说明臭氧可导致花生蛋白结构的部分展开或者发生蛋白聚集,从而使其严谨有序的二级结构趋于松散[33, 34]。此外,臭氧处理后蛋白二级结构的变化也可能与二硫键共价聚集有关,从而导致蛋白发生部分聚集,这与图1的结果相一致。
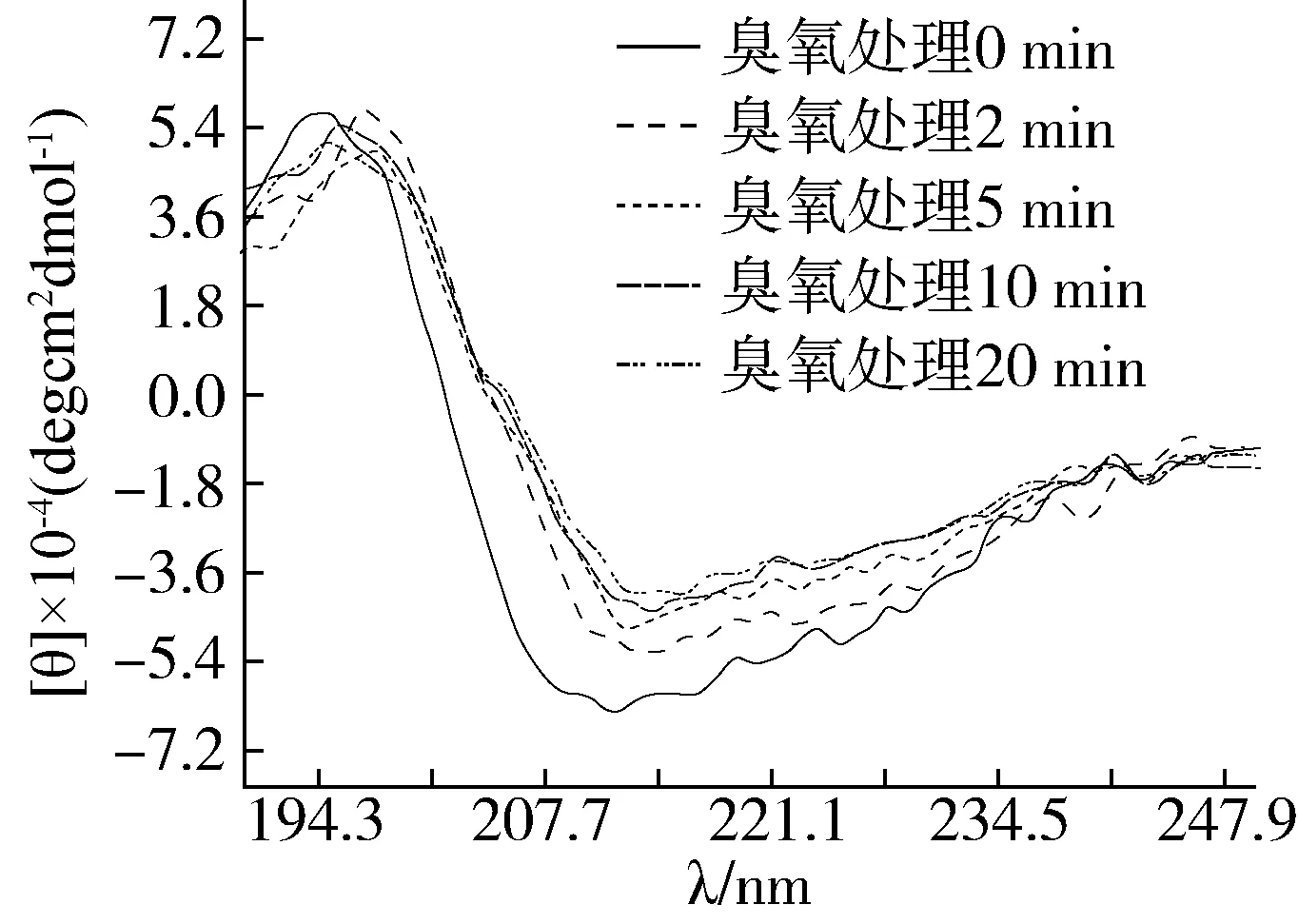
图4 臭氧处理前后花生蛋白的圆二色谱图

表3 臭氧处理前后花生蛋白的二级结构组成
2.7 臭氧处理花生蛋白的电泳分析
臭氧处理不同时间条件下花生蛋白的非还原和β-巯基乙醇还原型SDS-PAGE结果如图5所示。由图5a可以看出,大分子蛋白条带(>250 ku)发生明显迁移现象,随臭氧处理时间的延长,该条带亮度逐渐增加且上移。推测原因是臭氧处理使蛋白分子间的二硫键发生一定程度的聚集和交联,形成新的大分子蛋白聚集体[24, 35],这与二硫键含量随臭氧处理时间延长而增加的结果(图1)相符。而由图5b可知,>250 ku条带和花生球蛋白II亚基(61 ku)条带并未发生迁移但颜色明显变浅甚至消失,这可能是因为在还原态电泳中,β-巯基乙醇的加入打断了大分子蛋白聚集体中的二硫键。相比于非还原态电泳,经β-巯基乙醇处理后的花生球蛋白(40.5、37.5、35.5、23.5 ku)和伴花生球蛋白I亚基(18.0、17.0、15.5 ku)条带明显变粗变亮,但臭氧处理的影响效果甚微。由此可知,臭氧处理可诱导花生蛋白中二硫键发生一定程度的交联和聚集,其处理时间与聚集程度呈正相关,进而在一定程度上改变了蛋白结构。
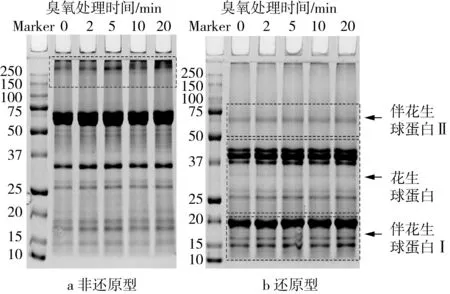
图5 臭氧处理前后花生蛋白的SDS-PAGE电泳图
3 结论
本研究借助氨基酸分析、圆二色谱、SDS-PAGE等技术深入表征臭氧处理对花生蛋白结构特性的影响。研究发现,臭氧处理可显著降低苯丙氨酸和含硫氨基酸(甲硫氨酸和半胱氨酸)含量,且其含量随处理时间延长而逐渐下降。相似地,随着臭氧处理时间的增加,花生蛋白的巯基含量显著下降,而二硫键以及羰基含量则呈上升趋势,这可能与臭氧促进蛋白质氧化有关。此外,臭氧处理有助于提升花生蛋白的热稳定性,其变性温度和吸热焓随臭氧处理时间(0~10 min)延长而增加,表面疏水性总体则呈下降趋势。有趣地是,臭氧作用可诱导花生蛋白中α-螺旋、β-折叠以及β-转角结构比例的下降,而无规则卷曲结构则呈现显著上升趋势,说明臭氧处理可导致蛋白质的二级结构发生明显变化。SDS-PAGE电泳结果也表明,臭氧氧化可诱导花生蛋白中二硫键发生部分交联和聚集,产生大分子蛋白聚集体(>250 ku)。综合而言,臭氧主要通过作用于分子内的巯基和二硫键而改变花生蛋白的结构特性。
