热休克蛋白70介导的表没食子儿茶素没食子酸酯抑制埃可病毒9型的活性研究
2021-02-04汪洋张辉马东波伍冬冬邓翔李芳郭胜存吴秋歌
汪洋 张辉 马东波 伍冬冬 邓翔 李芳 郭胜存 吴秋歌
郑州大学第一附属医院呼吸与危重症医学科(郑州450052)
埃可病毒(echovirus,ECHO)是一种小分子RNA 病毒,属于肠道病毒B 型,肠道病毒属,小核糖核酸病毒科,是全世界病毒性肺炎和无菌性脑膜炎最常见的病原体之一[1]。ECHO 分为34 型,各型致病力和致病类型也不同,其中6、9、19 型致病力较强。埃可病毒9 型(echovirus 9,ECHO9)是引起婴幼儿上、下呼吸道感染的常见病原体,病死率较高,在呼吸道疾病中的地位不容忽视[2]。临床表现复杂、多样化,症状轻重不等,从隐性的无症状感染、上呼吸道轻度不适到严重中枢神经系统感染、心力衰竭均可发生[2-3]。尽管目前已有中药成份的治疗埃可病毒感染的研究报道[4-5],但其机制研究较少,且至今仍然没有特异针对性的治疗措施或疫苗。目前临床上治疗埃可病毒感染主要应用更昔洛韦和干扰素,可结合中药辅助治疗,或给予对症支持治疗[6]。对于危重症患者,可静脉注射高效价免疫球蛋白,有利于减少危重症及死亡病例的发生。目前的治疗虽可减轻临床症状,缩短病程,但疗效尚不稳定,副作用多,治疗周期长,患者花费高。目前还没有开发出安全有效的预防疫苗。因此,开发高效、安全、副作用少的防治埃可病毒感染的新药,尤其是天然活性成分的化合物具有重大意义。
表没食子儿茶素没食子酸酯(epigallocatechin-3-gallate,EGCG)为茶多酚中主要的活性成份,约占总茶多酚含量的55%~65%[7]。研究表明,EGCG 具有抗氧化、抗肿瘤、抗细菌、抗炎、抗病毒、抗血栓形成及抑制血小板聚集的作用[8-10]。其抗病毒治疗的作用靶点及具体机制尚不明确。本研究旨在探讨EGCG体外抗ECHO9型的作用及机制,评价其作为天然药物治疗靶点的可行性。
1 材料与方法
1.1 材料ECHO9:为郑州大学第一附属医院临床分离株,来源于一名患者的脑脊液。该患者为32 岁男性,受凉1 d 后出现上呼吸道感染、高热,伴头痛、恶心、呕吐,自服解热镇痛药物3 d 后呼吸道症状、发热较前缓解,但头痛、呕吐较前加重。查体脑膜刺激征阳性,脑脊液检查未见白细胞增高及蛋白、糖和氯化物的改变,考虑病毒性脑膜炎。Vero 和Hep-2 细胞为本科室保存。EGCG 由Sigma公司提供,母液浓度为20 mg/mL,过滤除菌分装,4 ℃避光保存。Anti-Respiratory Syncytial Virus兔多克隆抗体(ab12253)和山羊抗兔IgG H&L(HRP)(ab6721)二抗购自美国Abcam 公司,抗GAPDH 兔多克隆抗体购自杭州贤至生物科技有限公司;小干扰RNA(siRNAs)(siControl,cat. no. sc-37007,序列为5′-AAUUCUCCGAACGUGUCACGU-3′;siHsp70,cat. no. sc-29352,序列为5′-CGGUGGUGCAGUCGGACAUGA-3′)和Hsp70 的RT-PCR 引物(h)-PR:sc-29352-PR购自Santa Cruz Biotechnology,Inc.。
1.2 方法
1.2.1 病毒的扩繁提前一天将Vero 细胞传代,使一瓶细胞扩繁为三瓶。次日细胞长至80%左右汇合度的细胞单层时,接入埃可病毒种子毒,置37 ℃、5% CO2培养箱中培养,观察细胞病变。待大部分细胞出现病变时,即由梭形变为圆形时,表明细胞已受病毒侵袭,部分凋亡坏死,收取细胞培养上清,以3 000 r/min 离心去除细胞碎片,将上清进行1 mL/管进行分装,置于-80 ℃冰箱保存。
1.2.2 病毒滴度测定在接毒前将生长状态良好的正常Vero 细胞铺96 孔细胞培养板。次日待细胞汇合度长至约80%时,弃去培养上清。将埃可病毒用细胞维持液从10-1至10-8做10 倍系列稀释,每个稀释度设6 个重复孔,将各稀释度的病毒液取100 μL接种于单层Vero细胞中,置37 ℃、5%CO2培养箱中培养,观察细胞病变程度并记录。采用Reed-Muench 法计算病毒培养上清和不同药物浓度处理细胞上清中的埃可病毒滴度。
1.2.3 EGCG的药物毒性测定将EGCG分别用细胞维持液稀释为400、200、100、50、25、12、6、3 μmol/L 8 个浓度,然后分别加入Vero 细胞已长成单层的96 孔细胞培养板中,每孔100 μL,每一浓度设置3 个复孔,同时设立正常细胞对照组,置37 ℃、5%CO2培养箱中培养,观察48 h。每孔加入CCK8 10 μL,避光培养1 ~4 h。酶标仪(波长450 nm)测定各孔光吸收值。各实验批次间重复6 次。
1.2.4 EGCG 的抗病毒效果检测已长成单层的
Vero 细胞以含不同浓度药物的维持液(各浓度均不超过半数致死量CC50)处理2 h 后,按照MOI(multiplicity of infection,MOI)=0.01 进行接毒,随后置于37 ℃、5 % CO2培养箱孵育2 h。弃去含病毒维持液,以无血清培养基洗1 遍,再更换含不同浓度药物的维持液,置于37 ℃、5 % CO2培养箱内培养。培养48 h 后,收集上清。测定病毒滴度,方法同1.2.2 所述。
1.2.5 Western blot 检测RSV 病毒蛋白的表达情况Vero 细胞铺于六孔细胞培养板,药物和溶剂DMSO 分别预处理细胞2 h,再以MOI 值0.01 接入埃可病毒,培养24 h 后,不同处理细胞样品进行电泳并转膜。NC 膜分别加入1∶1 000 稀释的抗GAPDH 兔多克隆抗体或抗埃可病毒F 蛋白抗体,再加入辣根过氧化物酶标记的羊抗兔IgG 二抗于37 ℃条件下摇床孵育1 h,化学发光系统成像拍照分析。
1.2.6 qPCR 检测病毒基因的抑制情况将埃可病毒的糖蛋白G 基因和猴GAPDH、Hsp70 基因分别录入在线qPCR 引物设计网站,设计qPCR 引物并进行合成,具体引物如下:G-F:5′-AAGCCAACCATCAACACCAC-3′,G-R:5′-TGGGCTTAGATAGCCTTCGG-3′;GAPDH-F:5′-TCGTGGAAGGACTCATGACC-3′,GAPDH-R:5′-CCATCCACAGTCTTCTGGGT-3′。将不同处理方法处理的六孔板细胞以MOI 0.01 接入埃可病毒,培养24 h 后,弃去上清,以PBS 洗细胞1 次,每孔加入1 mL Trizol 裂解,抽提细胞总RNA,并反转录为cDNA。PCR 运行程序为:95预变性10 min,随后进行40个循环:95 ℃10 s,56 ℃1 min 以及72 ℃10 s。采用SYBR Green 法进行扩增。
1.2.7 Hsp70的RNA干扰Hep-2细胞分别以Hsp70和对照的siRNA 进行转染,转染12 h 后,以MOI=0.01 接埃可病毒进行培养,分别在感染24、36、48 h收集培养上清,测病毒滴度。测定Hsp70 转录本的表达情况。具体参考产品说明书和方法1.2.2、1.2.6 进行。
1.2.8 统计学方法用SPSS 21.0软件统计分析,不同处理组的样品转录本、病毒滴度等采用t检验进行组间两两比较,检验水准α=0.05。
2 结果
2.1 病毒滴度测定结果取100 μL 病毒母液,接入长成单层细胞的T-25 细胞瓶中,每天观察细胞病变情况。当Vero 细胞中大部分出现病变时收毒(图1),测定病毒培养液中病毒滴度,记录每个稀释度不同孔的病变情况,经Reed-Muench 法算得每0.1 mL 母液中病毒滴度TCID50(半数组织培养感染剂量)为10-4.5,可以满足后续实验的需求。
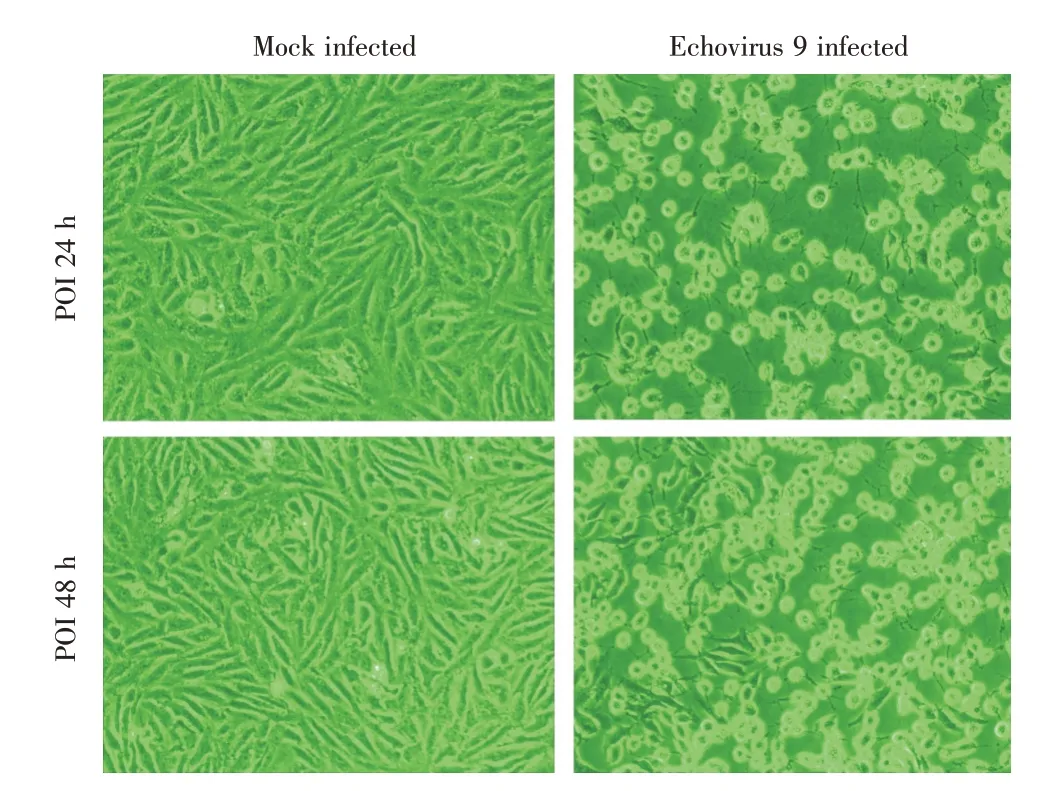
图1 ECHO 9 病毒在Vero 细胞上产生的病变(50×)Fig.1 ECHO 9 infected Vero cells
2.2 EGCG 的药物毒性测定结果不同浓度的EGCG 处理细胞48 h 后,测定细胞活力。EGCG 的细胞毒性具有明显的浓度梯度依赖反应,其中25 μmol/L 对细胞毒性与其以下浓度差异无统计学意义,因此,EGCG 对Vero 细胞的最大无毒浓度在25 μmol/L 以下(表1)。
2.3 EGCG 对ECHO9 病毒增殖的抑制效果为了更加直观清晰的评价EGCG 的抗RSV 效果,本研究采用测定上清病毒滴度TCID50的办法对药物进行药效学检测。EGCG 对细胞上清中释放病毒滴度的抑制结果如表2 所示,EGCG 分别在25 μmol/L和20 μmol/L 及以上浓度时,可抑制60%以上的病毒增殖;同时,在不同的EGCG 浓度作用下,培养上清中子代病毒释放量随着EGCG 浓度的增加而下降。结果显示EGCG 对ECHO9 具有体外抗病毒效果。
表1 不同EGCG 药物浓度下细胞活力Tab.1 Cell activity at different EGCG concentrations ±s
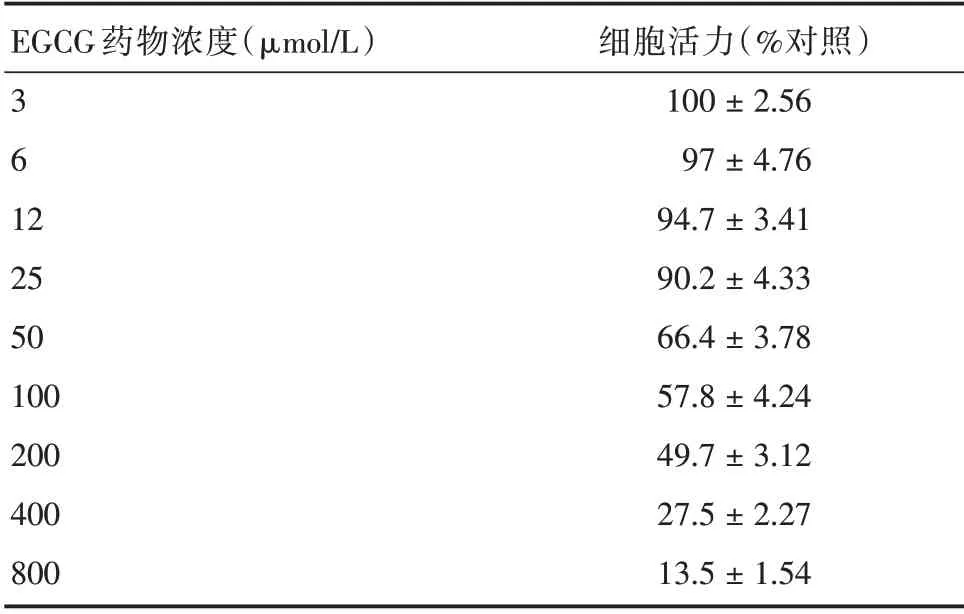
表1 不同EGCG 药物浓度下细胞活力Tab.1 Cell activity at different EGCG concentrations ±s
EGCG 药物浓度(μmol/L)361 2 25 50 100 200 400 800细胞活力(%对照)100±2.56 97±4.76 94.7±3.41 90.2±4.33 66.4±3.78 57.8±4.24 49.7±3.12 27.5±2.27 13.5±1.54

表2 不同浓度EGCG 对病毒增殖的抑制作用Tab.2 Inhibition effect of EGCG at different concentrations on virus
2.4 Western blot 检测ECHO 9 病毒蛋白表达受EGCG 抑制的情况将不同处理方法(正常细胞、溶剂DMSO 和EGCG 12 μmol/L)处理的细胞样品的检测结果如图2 所示,在膜上显现出相对分子质量48 kD的条带,这与ECHO 9病毒G蛋白分子量大小一致。在EGCG 处理的样品中,在细胞内参蛋白GAPDH 含量相同的情况下,病毒蛋白表达量显著低于对照组和溶剂处理组的细胞样品(Western blot结果分析相对表达量分别为0.98、0.96、0.12),表明EGCG 使病毒在细胞中的增殖受到明显抑制。
2.5 qPCR 检测EGCG 处理细胞中病毒基因的转录抑制情况由图3A 可知,本研究所设计引物具有单一的溶解曲线峰,说明扩增效果良好。由于病毒mRNA 合成在病毒感染后进行,比较了药物处理和未处理的细胞在感染24 h 后病毒G 蛋白mRNA 的转录水平。EGCG(12 μmol/L)、DMSO 溶剂和ECHO 9 病毒感染等不同处理的细胞中病毒G 基因的转录水平以GAPDH 进行归一化处理,并按照ΔΔCt 方法进行计算。实验结果(图3B)显示,EGCG 的处理使ECHO9 病毒糖蛋白G 基因组转录下降了约85.3%,与溶剂组和正常对照组相比差异有统计学意义(P<0.01)。
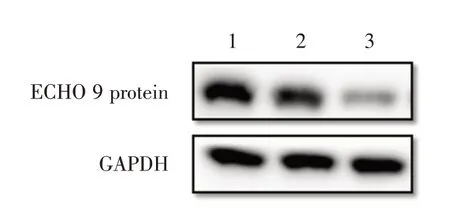
图2 Western blot 检测ECHO 9 病毒蛋白表达Fig.2 ECHO 9 protein expression by Western blot
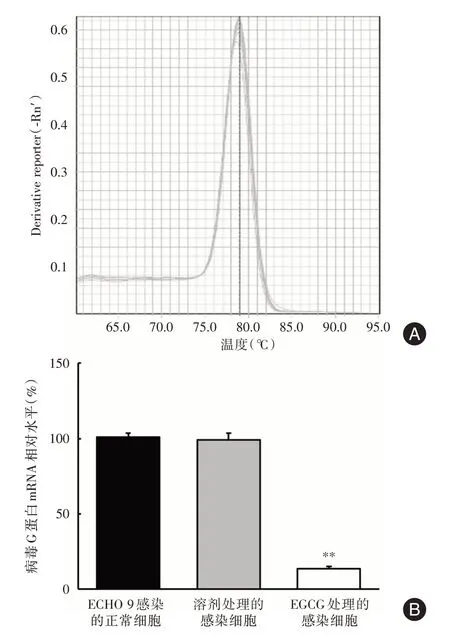
图3 不同处理对ECHO9 G 蛋白基因转录水平的影响Fig.3 Effect of different treatments on ECHO9 G mRNA expression
2.6 EGCG处理对Hsp70的转录和表达的影响为了进一步探讨EGCG 抑制ECHO9 病毒的作用机制,对EGCG 处理的细胞中Hsp70 和糖蛋白G 基因的转录本进行实时荧光定量PCR 检测。EGCG 的处理能够使Hsp70 的转录本水平下调约60%,在病毒感染细胞能够达到相同的效果;而病毒ECHO 9的感染没有使Hsp70 转录水平发生明显的变化(P>0.05,图4A)。同时,在蛋白水平上,EGCG的处理使得ECHO 9 感染前后的细胞中的HSP70 的表达水平受到明显抑制(P<0.01,图4B),其蛋白相对表达量分别为1.36、0.71、1.42 和0.63。
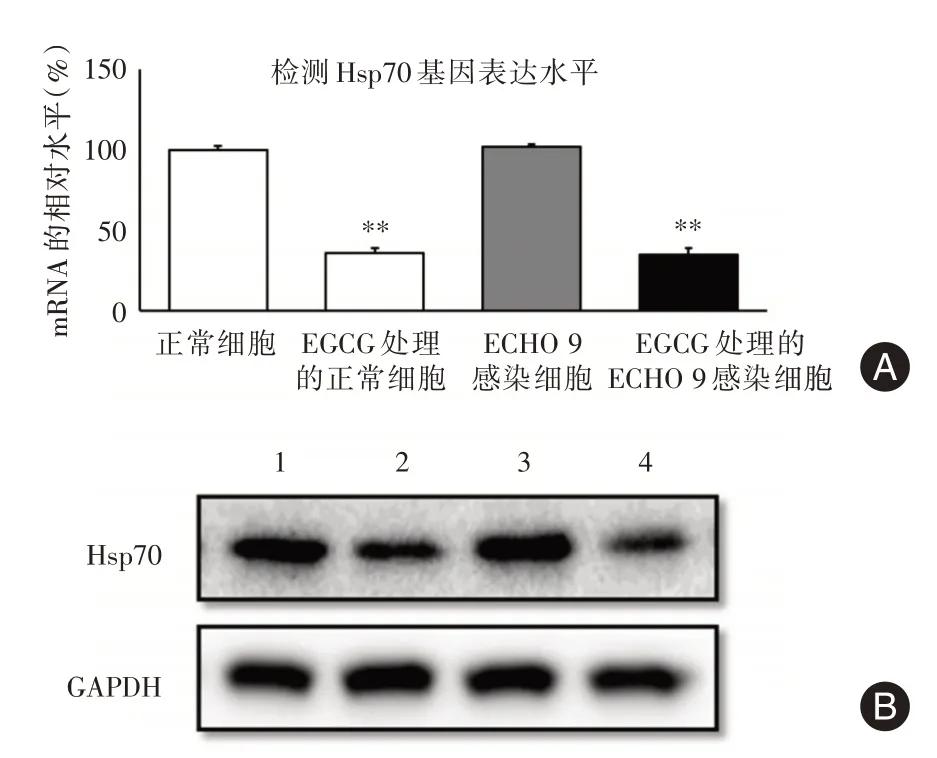
图4 EGCG 的处理前后ECHO 9 感染前后细胞中的Hsp70的表达水平Fig.4 Hsp70 expression level of ECHO 9 infected cells before and after EGCG treatment
2.7 RNAi 证实Hsp70 为ECHO 9 的治疗靶标鉴于EGCG具有多种生理活性,但其抑制ECHO 9的增殖是否是通过抑制其特异性底物Hsp70 来实现有待研究。本实验采用RNA 干扰的手段进行了初步探索。RNA 干扰敲低了细胞中HSP70 的表达水平。经Western blot验证siHsp70干扰效率为89.7%,干扰效果良好。不同处理组Hep-2 细胞培养上清中病毒的一步生长曲线显示,细胞培养上清液中病毒滴度siHsp70 组(102.03TCID50/0.1 mL)显著低于实验对照组(104.98TCID50/0.1 mL)(P<0.01,图5)。
3 讨论
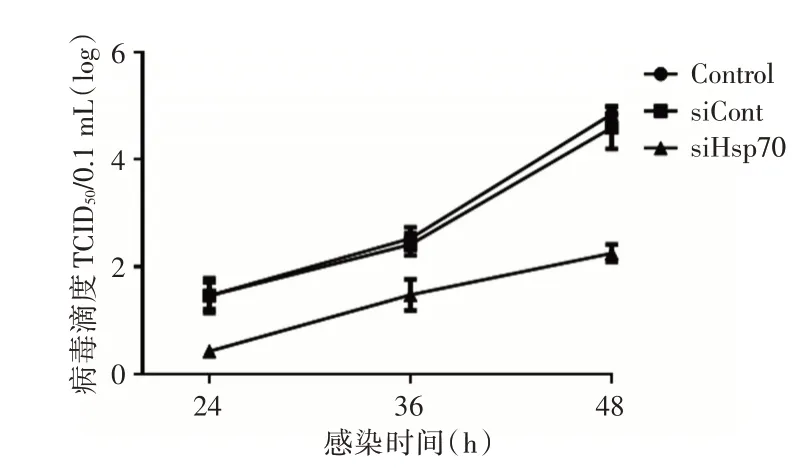
图5 不同处理Hep-2 细胞培养上清中病毒的一步生长曲线Fig.5 One step growth curve of virus in supernatants with different treatments of Hep-2 cells
ECHO 是全世界病毒性肺炎和无菌性脑膜炎最常见的病原体之一,多发于夏、秋季,绝大多数是隐性感染。埃可病毒主要经粪-口途径传播,也可通过咽喉分泌物排出病毒经呼吸道传播。随着各国卫生条件的改善和医疗水平的提高,呼吸道传播逐渐成为首要途径。病毒从上呼吸道或口腔进入体内,在咽喉部或肠壁淋巴组织中停留并增殖,从原发部位向局部淋巴组织或血液转移并经血型播散带到其他器官如心脏、肝脏、脑、肾脏、骨骼肌、甚至皮肤黏膜,几乎所有细胞均能受累[11-12]。埃可病毒9 型(Echovirus 9)是从无菌性脑炎患者检测到的最常见类型,散发或流行均可出现[12-13]。其感染的临床表现类似于风疹,对新生儿尤其具有破坏性,可诱发重症肺炎、肝功能衰竭、脑膜炎及脑炎,脓毒血症,最终导致多脏器衰竭甚至死亡[2,13]。
茶多酚是从茶叶中提取的多羟基酚类混合物,其中以儿茶素占茶多酚总量的60%~80%,包括表没食子儿茶素没食子酸酯(EGCG)、表没食子儿茶素(EGC)、表儿茶素没食子酸酯(ECG)、表儿茶素(EC)[14-15]。EGCG 是茶叶中特有的儿茶素,含量最高,约占茶多酚制品的50%左右[7]。研究表明EGCG 具有抗氧化、抗感染、抑制肿瘤生长、提高免疫力,且对痢疾、伤寒、金黄色葡萄球菌等微生物具有明显的抑制作用[21]。体外实验已证明EGCG 可通过抑制HIV-1 逆转录酶的活性发挥抗HIV 作用[22]。EGCG 还可通过抑制细胞内内涵体或溶酶体的酸化来降低流感病毒的感染率[23]。目前尚无EGCG 抑制埃可病毒的相关文献。
热休克蛋白(heat shock proteins,HSPs),是从细菌到哺乳动物中广泛存在的类热应急蛋白质。当有机体暴露于高温环境会激发合成此种蛋白,以保护机体,许多热休克蛋白具有分子伴侣活性。按照蛋白大小,热休克蛋白共分为五类,分别为Hsp100、Hsp90、Hsp70、Hsp60 以及小分子热休克蛋白[19]。其中Hsp70 通过活化天然免疫细胞的方式参与天然免疫,通过调节抗原呈递细胞对抗原的加工与呈递,增强免疫应答,参与免疫球蛋白组装,发挥对适应性免疫的影响,进而参与感染与免疫[20]。GAIE 等[21]研究发现,随着感染的发生Hsp70 会被转运至病毒诱导包涵体中,与病毒N 蛋白和脂筏共定位。前期的研究[22]发现,Hsp70 在埃可病毒感染中发挥一定的功能,可能是抑制埃可病毒的治疗靶点。
本研究中,研究了EGCG 的体外抑制ECHO 9的作用,证实其作为抗病毒药物的治疗潜力。EGCG 能使Hsp70 的转录本水平下调,在病毒感染细胞中亦能达到相同的效果;而病毒ECHO 9 的感染没有使Hsp70 转录水平发生明显的变化,这与之前的研究结果一致[22]。同时,在蛋白水平上,EGCG 的处理使得ECHO 9 感染前后的细胞中的Hsp70 的表达水平受到明显抑制。因此,本研究推测,EGCG 具有良好抗ECHO 9 效果是通过抑制Hsp70 的蛋白表达,进而影响病毒感染中诱导的内涵体和脂筏的形成,从而使病毒子代的产生受到影响。因此,本研究进一步通过qPCR、Western blot 和商业化RNA 干扰序列的转染成功验证了设想,即EGCG 可能是通过作用于Hsp70 实现抑制ECHO 9 病毒的增殖的。本研究中采用了直接测定药物处理后细胞上清中病毒滴度的办法测定药物抗病毒效果。同时,结合细胞中病毒蛋白表达情况和基因转录情况,确认了EGCG 的抗ECHO 9效果及其初步的作用机制。
综上所述,EGCG 在体外能有效抑制ECHO 9病毒,其作用机制可能是通过下调Hsp70 实现的。作为一种有效的天然茶多酚成分,有望用于埃可病毒感染引起的呼吸道疾病的预防与治疗。
