一个大麦突变体库的构建及其叶宽突变体的细胞学初步分析
2021-01-28秦丹丹许甫超彭严春葛双桃焦春海
秦丹丹 许甫超 徐 晴 彭严春 葛双桃 董 静 焦春海
(1湖北省农业科学院粮食作物研究所,湖北 武汉 430064;2 粮食作物种质创新与遗传改良湖北省重点实验室,湖北武汉 430064)
突变体的创制、筛选和利用可以为种质创新和育种提供有价值的基础材料。上世纪20年代,美国科学家首次提出了通过离子辐射进行诱变可以增加大麦的突变频率,而且所获得的突变可以稳定遗传给后代[1],开启了植物研究的新领域。
通过理化手段进行诱变从而创制新的种质资源,可以提高基因交换和基因重组的频率,并打破不利基因连锁[2],创造丰富的遗传变异,拓宽原品种的遗传基础,缓解亲本资源匮乏、遗传基础狭窄等问题,促进品种遗传改良和突破性品种的培育[3-4]。根据联合国粮农组织和国际原子能机构(FAO/IAEA)的数据库,截止2020年10月24日,已有超过3 362 个作物新品种是通过诱变育种方法获得的,其中80%左右是由物理诱变获得的[5]。我国辐射诱变育种研究始于1956年,在培育新品种和创制新种质方面发挥了重要作用[6]。据不完全统计,截止2018年8月,我国利用诱变技术直接或间接育成和审定了1 033 个植物新品种,位列全球第一位,创造了巨大的社会经济效益。
随着拟南芥、水稻等模式植物以及其他植物基因组测序的完成,植物的功能基因组学研究进入了飞速发展阶段。通过产量、品质、抗逆性等相关突变体进行调控基因的克隆则是获得相关功能基因的重要途径,如水稻矮秆基因Dwarf[7]、雄性不育基因MER3[8]、粒长基因GLA1[9],玉米株型调控基因ZmACS7[10],二倍体小麦的脆秆基因TmBr1[11],小麦叶绿素和脂肪酸合成相关的镁螯合酶基因等[12],都是利用突变体克隆获得的。这些研究为解析相关性状形成的分子机理并通过分子育种的手段进行目标性状的遗传改良奠定了基础。
大麦(Hordeum vulgareL.)是全球也是我国第四大谷类作物,具有食用、饲用和酿造等多种用途。大麦一直是我国青藏高原地区最具优势的粮食作物之一[13]。随着社会的发展和人们生活水平的提高,大麦作为功能型食品,在预防和治疗高血糖、高血压等慢性病中的作用也逐渐被大众认知和接受[14]。但目前我国的大麦生产存在自给率低、对外依存度高等问题。根据海关数据,2019年1—11月,我国从加拿大、澳大利亚等大麦主要出口国进口大麦596 万t。因此,培育高产专用型大麦品种对加快我国畜牧业产业化进程并改变大麦依赖进口的局面,以及保障我国粮食安全具有重要意义。此外,大麦还具有早熟、耐寒、耐旱、耐盐碱、耐瘠薄等特性,也是作物进行遗传改良的重要基因来源[15]。大麦和小麦在基因组成及排列顺序上存在较高的保守性,且二者在农艺、产量和品质等性状方面也具有很高的相似性,但基因组只有小麦的1/3,因此在基础研究中,大麦常被作为小麦的模式植物进行研究[16]。
目前,通过甲基磺酸乙酯(ethyl methyl sulfonate,EMS)诱变构建了二棱大麦Optic[17]和Barke[18]的突变群体,利用叠氮化钠构建了Morex[19]和Lux[20]的突变体库。我国科学家利用EMS 构建了自育大麦品种浙农大3 号[21]和国外引进种质Tamalpais[22]的突变体库。突变体在大麦重要功能基因的克隆中也发挥了举足轻重的作用,如大麦淀粉合成相关基因Fra[23]、分蘖和叶形调控基因HvHNT1[24]等,都是利用突变体通过图位克隆的方法获得的。通过诱变处理获得的特殊种质,为克隆目标性状调控基因奠定了基础,如从北青7号突变体库中筛选的穗分枝突变体[25],从浙农大3 号中筛选的蜡粉缺失突变体P1[26]等。但是,上述突变体库均采用单一诱变方法,所获得的突变类型有限,而且利用自育大麦品种构建突变体库的报道相对较少。
为构建具有自主知识产权的大麦突变体库,本研究通过60Co-γ 射线辐射和EMS 复合诱变,构建了自育品种鄂大麦934 的突变体库,并对叶宽突变体的叶片细胞进行显微观察,旨在为大麦种质资源创新以及重要性状调控基因的克隆和形成机理解析提供材料基础。
1 材料与方法
1.1 供试材料
鄂大麦934 是由湖北省农业科学院粮食作物研究所选育的粮草兼用型大麦品种,于2014年通过湖北省品种审(认)定(鄂审麦2014003),并于2018年进行了国家非主要农作物品种登记[GPD 大麦(青稞)(2018)420016],品种权号CNA20151702.8。根据湖北省大麦区域试验结果,鄂大麦934 属二棱皮大麦,株型较紧凑,幼苗半匍匐生长,分蘖力较强,生育期184 d,株高80.8 cm,穗粒数24.6 粒,千粒重39.3 g。
1.2 材料种植及处理方法
1.2.1 材料种植 本试验所有材料均种植于湖北省农业科学院粮食作物研究所核心试验田(湖北,武昌)。材料的田间种植、性状记载及成熟后收获均在大麦正常生长季内完成,田间栽培管理措施同当地、当季常规种植大麦。
1.2.2 材料处理 2012年夏正常收获的鄂大麦934种子,及时晾干和脱粒。挑选大小均匀一致、健康无病害的鄂大麦934 饱满种子1.5 kg。首先利用总剂量为320 Gy 的60Co-γ 射线辐射处理39 min(剂量率为1.92~26.00 Gy·min-1)(湖北省农业科学院辐照中心)。然后将处理后的种子在室温下浸泡1 h 后用1%EMS(v/v,美国Sigma 公司)溶液浸泡,并置于摇床上轻微晃动,确保溶液和种子充分接触。处理8 h 后用自来水冲洗3 h,除去胚上残留的EMS,并在通风处进行短时间晾干。
1.2.3 发芽率和发芽势 统计经诱变处理和未经处理(对照)的种子各100 粒,均匀摆放在铺有湿润滤纸的培养皿中,置于恒温恒湿培养箱中,温度设置为22℃/18℃,光照时间为12 h。培养期间及时补充水分,保证滤纸处于湿润状态,7 d 后统计发芽率、苗高、根数和主根长。
1.3 突变体鉴定
取诱变处理后的M0种子15 000 粒进行点播,行长3.0 m,行距25 cm,株距5 cm,每50 行种植1 行未经处理的鄂大麦934 作为对照。成活M1植株进行性状初步调查,挑选与对照表现不同的单株按单株收获,剩余单株收获单穗。将收获的单株和单穗,每株或每穗取1 粒种子混合点播成M2群体,种植方法同上。对成活单株进行全生育期调查,选择在株高、株型、生育期、叶型、叶色、穗长、穗度、穗型、粒形、结实率等性状中至少有一个与对照表现不同的单株,挂牌并及时收获、晾晒和保存。将M2收获的单株种植成株行并调查,选择至少有一个性状与对照表现不同的株行,从中挑选典型单株挂牌和收获,行内表现为分离的株行则分别挂牌和收获。对收获单株继续种植,并对目标性状提纯,直至获得行内纯合的株行,按行收获。
1.4 叶片细胞显微结构观察
挑选叶宽与野生型相比有显著差异的突变体3 个(叶细1 个,叶宽2 个),与野生型一起,剪取抽穗期旗叶,通过番红-固绿法观察并比较叶片细胞大小和形态等变化。该试验由武汉谷歌生物科技有限公司协助完成,具体步骤参照公司说明书。
2 结果与分析
2.1 复合诱变后种子的发芽率及发芽势
为了明确所实施的60Co-γ 和EMS 复合诱变处理对大麦种子活力的影响,对经处理和未经处理(对照)的鄂大麦934 种子7 d 发芽率及生长势进行了分析。结果表明,经处理的种子发芽率显著降低,幼苗和主根生长明显受到抑制,但是根数变化不大(表1)。

表1 诱变处理后种子性状测定Table 1 Seed traits of control and treatment
2.2 突变体库性状调查
将处理后的15 000 粒M0种子种植后,除诱变处理对种子本身发芽率产生了较大影响外,部分出苗后的植株因为白化或者其他原因死亡,存活并最终可正常收获的M1植株共8 432 株。存活植株中,有部分在M1即表现出与对照不同的性状。对这些变异单株挂牌、统计和收获,共计收获405 株(部分植株可能表现为多重性状)。其中涉及株高变异的67 株、株型14株、生育期50 株、叶片大小36 株、叶片形状8 株、叶片颜色10 株、穗宽12 株、穗长43 株、穗分枝形态4 株、粒形60 株、育性降低120 株。其他单株每株收获1穗。从以上收获的单株和单穗上各取1 粒种子混合播种,并进行多轮筛选,最终获得与对照存在明显差异且性状无分离、可遗传的变异株982 株,性状涉及叶片、茎秆、穗部、籽粒、生育期和结实率等。有些突变体表现为相同或相似的性状,也有些突变体表现为多重性状突变。
2.2.1 叶片性状突变体 本研究所获得的叶片性状变异主要涉及叶片形状、叶片颜色以及叶片与茎秆夹角。叶片形状变异包括叶长、叶宽、叶厚、圆叶、卷叶、早衰、皱缩等(图1)。野生型鄂大麦934 抽穗期的旗叶平均长度为15.9 cm,宽度为1.4 cm。对突变群体中各材料进行测量发现,旗叶长度变异在8.64 ~23.30 cm 之间,宽度变异在0.56~2.20 cm 之间(图1-b)。
叶片颜色变异主要表现为叶色深、叶色浅、叶片金黄色、白色、条纹白、灰绿色、锈状斑点、烫伤状枯死等(图1-c~f)。这些叶片颜色变异通常从苗期即开始表现,贯穿整个生育周期,有的则可以在后期转为正常叶色,如白色叶片突变体。
根据叶片与茎秆夹角是大还是小,相应的表现即为叶披或者叶挺。本研究发现了两种极端类型的叶夹角突变体,其中一种旗叶较为直立,与茎秆夹角约为20°(图1-g),另外一种旗叶在穗子完全抽出前即已下垂,与茎秆的夹角接近180°(图1-h)。
其他类型的叶片变异还包括叶片早衰,并鉴定出一种特殊的叶片变异,表现为旗叶正常,但是穗子基部还包围着一层苞叶状叶片(图3-i)。
2.2.2 茎秆性状突变体 茎秆性状涉及株高、茎粗细、茎秆颜色、茎秆节数、茎秆蜡质、分蘖、株型等(图2)。本研究中,鄂大麦934 的平均株高为84 cm,所发现的茎秆长度突变体株高变异范围较广,在30 ~103 cm 之间(图2-a)。此外还发现一种表现为茎秆节数增多但株高变矮的突变类型,该突变体茎秆平均节数可达10 节,较野生型的5 节显著(P<0.05)增多,但是株高只有40 cm,不足野生型的50%(2-b)。
蜡质合成与植物的抗逆性密切相关,本研究发现的蜡质合成突变体,茎秆表现为完全无蜡,呈翠绿色(图2-c)。2019—2020年度田间观察发现,该突变体叶片和茎秆上出现了严重的、不同于锈病的锈状斑点,但正季播种的同一突变体未观察到这一性状。在所鉴定的突变体中,还有一株表现为茎秆紫色(图2-d)。
野生型的鄂大麦934 具有很强的分蘖能力,可以形成10 个以上的分蘖。在包含有982 个突变体的群体中,共检测到43 株表现为分蘖增多(图2-e),分蘖可超过50 个。但多数分蘖增多的突变体,还伴随着茎秆变细、穗子变小、千粒重降低等性状。此外,还有植株表现为分蘖变少,只能形成1~3 个分蘖。
株型一直是大麦育种家所关注的重要性状之一,本研究也发现了株型发生改变的突变类型,包括紧凑型、分散型和丛生等(图2-f、g)。
此外,抗倒性是大麦极为重要的育种目标之一,茎秆粗度通常被认为是影响植株抗倒伏能力的重要因素。成熟的鄂大麦934 茎秆直径平均为3 mm,本研究发现的茎秆粗细突变体中,茎秆直径变异在2 ~4 mm之间(图2-h)。

图1 叶片颜色和形态突变体Fig.1 Leaf color and morphology mutation

图2 茎秆性状突变Fig.2 Stem traits mutation
2.2.3 穗部性状突变体 在所鉴定的突变体中,穗长和穗宽均表现出较大变异。鄂大麦934 的平均穗长为8.5 cm(图3-a、b),突变群体中,穗长(不含芒)变异在5.3~10.4 cm 之间(图3-b)。芒是野生大麦种子得以在大自然中传播的重要器官,也是生长后期植株进行光合作用的场所之一。本研究所鉴定的突变体中,芒长也表现出较大变异。其中,鄂大麦934 为长芒大麦,但在突变群体中发现了短芒、无芒等变异类型(图3-c、d);还有一个突变株系顶部小穗为白色且不育,顶芒也为白色(3-e),且该性状稳定可遗传。
鄂大麦934 为二棱大麦,本研究鉴定出了多种其他棱型的突变体,包括侧小穗部分退化成圆点状、分枝穗、绒状花穗等(图3-f、g、h)。此外,上述提到的多节矮化突变穗部性状也与野生型不同,表现为同一分蘖上的多个节上均着生有穗子,而且最顶端穗子的基部还可以分化出多个小穗(图3-j)。此外,还发现一个突变体株系开花期花药外露(图3-k),以及一株穗柄扭曲(图3-l)。

图3 穗部性状突变体Fig.3 Spike morphology mutation
2.2.4 籽粒性状突变体 本研究所发现的籽粒性状变异主要表现在籽粒大小、籽粒形状和籽粒饱满度方面(图4),还有些突变体籽粒表现为黑胚。其中,籽粒大小突变体主要表现为籽粒变短(图4-a)或者变细,籽粒形状突变体主要表现为籽粒变圆(图4-b)。
2.2.5 其他发育性状突变体 根据湖北省大麦区域试验结果,鄂大麦934 生育期为184 d。本研究发现的生育期发生改变的突变体中,抽穗期最早和最晚的分别较同期种植的鄂大麦934 早和晚约20 d。
此外,本研究还发现了雄性不育和结实率下降的突变体(图5-a),该雄性不育突变体颖壳蓬松发亮,花药发育异常(图5-b)。
2.3 叶宽突变体叶片细胞显微观察
叶片是植物进行光合作用最重要的器官,在一定程度上决定了植物的最终产量。为了进一步分析本研究所获得的叶宽突变体形成的原因,分别挑选了1 个叶宽变细和2 个叶宽变宽的突变体,观察旗叶的叶片细胞并与同期野生型叶片(图6-a)进行比较。结果发现,这些叶宽突变体在细胞水平上表现各不相同。叶细突变体(图6-b)上表皮细胞呈现不规则状,且相邻叶脉之间的距离较野生型变大,而叶宽突变体(图6-c)相邻叶脉间的细胞数目与野生型相同,但细胞变小,因此导致相邻叶脉间的距离变短,另一叶宽突变体(图6-d)上表皮细胞较野生型变大,但相邻叶脉间的细胞数目减少,最终导致相邻叶脉间距离与野生型差异不明显。
3 讨论
构建有效的突变体库不仅是鉴定基因功能并进一步解析基因调控网络的有效途径,而且对作物品种遗传改良及拓宽遗传基础具有重要意义。获得突变体的方法包括自发突变、理化诱变和插入突变。由于自发诱变的通量有限,而插入突变对转基因技术的依赖性较强,因此理化诱变成为应用最广泛的方法。目前,包括大麦在内的突变库构建大多是采用单一的物理或者化学诱变方法。如通过EMS 诱变所构建的浙农大3号突变体库,共包含81 株稳定可遗传的突变株系,表型变异率约为7.46%[21];通过不同剂量7Li 离子束诱变获得的粒型、品质等变异株[27];也有少数通过理化复合处理对小麦[28]和大麦[29]进行诱变的报道。这些研究均发现复合处理对植株的影响大于单独处理。在大麦中,不同基因型所能承受的诱变强度差异较大,如利用总剂量为1 500 Gy(剂量率1 Gy·min-1)60Co-γ 处理浙秀2 号,发芽率仍可达70.3%,较对照(83.1%)只减少12.8 个百分点[30],而仅300 Gy(6.25 Gy·min-1)的60Co-γ 处理对昆仑15 号的株高、茎粗等性状就可产生显著影响[31]。为了最大限度地对大麦基因进行突变,构建具有自主知识产权的大麦饱和突变体库,本研究选用自育品种鄂大麦934,利用总剂量320 Gy的60Co-γ 和1% EMS 对其进行复合处理。

图4 籽粒形状突变体Fig.4 Seed shape mutation

图5 雄性不育突变Fig.5 Male sterile mutation
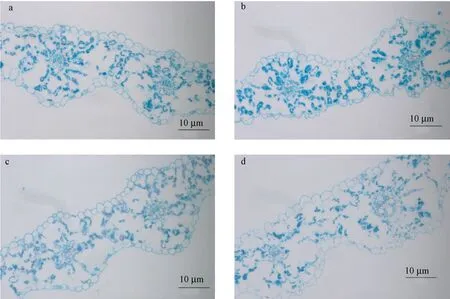
图6 叶宽突变体的细胞显微观察Fig.6 Microscopic analysis of cells of leaf width mutation
本试验结果表明,复合处理引起鄂大麦934 发芽率降低了43 个百分点,说明本研究中所采取的320 Gy的60Co-γ 射线和8 h 的1% EMS 为该品种半致死剂量,可以作为进行大麦复合诱变处理的参考剂量。在筛选过程中发现,有些突变株系在早代出现了性状分离,对这些单株分别进行了收获和统计,但由于未对根系、品质、抗逆性等性状进行考察,故没有对突变率进行统计。最终筛选出982 株在叶片、茎秆、穗部、籽粒、生育期、育性等性状发生了明显可稳定遗传的突变株系。
本研究所鉴定的突变类型中,有些性状研究较多,如叶色浅、分蘖性、株型、分枝穗和蜡质合成[16,23,25,32]等,虽然所用材料不同,但在后续研究中,仍需对相关突变体进行等位性检测,排除与其他研究重复或者相似的突变株系。有些性状则研究较少,如一蘖多穗、点状侧小穗等(图3)。此外,有些自然界中存在的性状,或者其他研究中所出现的变异类型,在本研究中未出现,如六棱大麦[33]、粒色变异[34]、皮裸性变异[16]等。鄂大麦934 为二棱大麦,在突变体库中,没有发现六棱的突变株系,但是发现了其他不规则棱形的突变体(图3)。这些性状的调控基因可能为已报道的棱形调控基因的等位基因,也有可能是新的基因,需要进一步研究明确。
通过多年观察发现,本研究所鉴定的蜡质合成突变体在不同的环境下种植出现了新的性状,初步推测该性状可能与环境湿度有关。这种现象在其他类型的突变体中也有报道,如通过X 射线诱变获得的大麦突变体mnd,在田间种植时表现为茎秆节数增多、株高变矮,但是在温室种植时表现正常[35]。因此,为了全面认识所鉴定的突变体,还需要对重点突变体进行多年多点和多环境观察,为深入解析这些性状形成的机理奠定基础。
高生物产量一直是包括大麦在内的饲草品种选育的重要目标之一,大叶是增加生物产量的有效途径[36]。本研究所构建的突变群体中,叶长和叶宽变异丰富(图1-b)。了解它们形成的原因是应用这些性状的基础。本研究对所鉴定的叶宽突变体的旗叶进行了细胞观察,发现不同类型甚至相同类型的突变体在细胞水平上表现各不相同,如叶细突变体的异常不仅体现在细胞形状的异常,而且相邻叶脉间的间隔也与野生型不同。同为叶片变宽的突变体,有的细胞大小没有发生改变,有的细胞大小则发生了改变。这些叶宽突变体在细胞水平上表现差异暗示这些性状可能由不同基因调控。如在大麦中,棱形调控基因VRS1 突变导致的叶片变宽是由叶脉数目增加引起的[37],而PHOTOPERIOD-H1 则主要通过影响细胞数目来调控叶片大小[38]。
多数研究认为通过理化诱变获得的突变多为隐性突变,M1所获得的变异在M2中不能稳定存在[39],所以在M1通常不进行选择。但是本研究在M1即收获了405 株初选突变体,在这些初选突变体中,约1/3 表现为高度不结实,还有的在下季种植过程中与对照无显著差异,最终保留982 株,其中约10%来源于405 株初选突变体。这说明通过理化诱变也可以获得显性突变,这种现象在小麦中也有报道[40]。
通常认为物理诱变会引起DNA 片段丢失、异位或者倒位,而EMS 化学诱变往往产生点突变。本研究所鉴定的突变体是通过理化复合诱变产生的,因此不能确定所表现出来的性状变异是由一种还是两种诱变方式引起的。但是,本研究利用几个突变体构建了分离群体,性状涉及叶色、棱型和茎秆节数,结果发现,这几个性状均由单隐性基因调控。如金黄叶色(图1-c)性状,利用集团分离分析(bulked segregant analysis,BSA)法成功将该基因定位在大麦基因组上约12.7 cM 的区间内,为该基因的精细定位和克隆奠定了基础[41]。在其他研究中也有通过物理诱变获得单基因突变体的报道,如通过γ 射线处理大麦品种Vlamingh所获得的多分蘖突变体[24]和经60Co 辐射诱变所获得的弯秆矮生性状调控基因[42],都是由单隐性基因调控的质量性状。
构建突变体库的最终目的是利用这些突变体及其调控基因进行新品种的培育,并服务于农业生产。如上世纪60年代,捷克利用核辐射诱变育成的矮秆高产大麦突变品种Diamant,不仅被直接推广利用至欧洲各国,而且被作为核心亲本加以利用,先后培育出150 余个具有Diamant 血统的优良品种并推广至欧洲、北美和亚洲各国,种植面积占全欧洲大麦面积的54.6%[43]。本研究所鉴定的直立株型、粗茎秆型、大叶型、大穗型、大粒型等包含有利性状的突变体,都可作为育种过程中的重要亲本资源加以应用。而对这些性状的调控基因进行克隆和功能鉴定也将是未来工作的重点。此外,还需对各突变体的根部性状、品质性状以及抗逆性等进行评价,从而筛选优异种质并加以利用。
4 结论
本研究采用60Co-γ 和EMS 复合诱变方法构建了自育大麦品种鄂大麦934 的突变体库,共鉴定了982株性状稳定且可遗传的突变株系,突变类型丰富,涉及叶片、茎秆、穗部、籽粒、发育等多个性状。对叶宽突变体的旗叶细胞大小进行显微观察发现,与野生型相比,各株系的细胞大小和形态呈现各不相同的变异类型。本研究结果为大麦功能基因组学研究及新品种选育提供了重要资源。
