大菱鲆对脂质及碳水化合物的需求研究进展
2021-01-27作者王卓铎孙广文张海涛
◆作者:王卓铎 孙广文 张海涛 刘 敏
◆单位:广东恒兴饲料实业股份有限公司;农业农村部华南水产与畜禽饲料重点实验室
大菱鲆(Scophthalmus maximus)又名多宝鱼,属鲆科(Bothidae)菱鲆属(Scophthalmus),生长迅速、营养丰富、经济价值高,是我国北方重要的肉食性海水养殖鱼类。脂质在大菱鲆生命代谢过程中具有多种生理功能,饲料中的脂肪更是鱼类必需脂肪酸的来源,是鱼类重要的营养成分之一。鱼类对糖类的利用率较低,且已有的研究证实大菱鲆幼鱼对饲料碳水化合物的利用能力较弱,当饲料糖含量超过15%时就会抑制幼鱼的生长,并会对内脏器官产生不良的影响(李晓宁,2011)。但饲料不含糖的情况下,鱼类将分解更多的蛋白质和脂肪来提供能量及合成生命所必需的其他物质。因此,基于饲料成本与环境保护考虑,使饲料含有适量的糖类也非常重要。
1 大菱鲆脂肪营养
脂类是鱼类生长所必需的一类营养物质,在鱼类生命代谢过程中具有多种生理功能,包括作为组织细胞的组成成分、提供能量、利于脂溶性维生素的吸收运输、提供必需脂肪酸、合成维生素与激素、提高饲料蛋白质利用率等(麦康森,2017)。鱼类的脂质需求即是其对能量和很多特殊功能脂质组分的总需求(麦康森,2015)。
1.1 脂肪
饲料中脂肪含量不足或缺乏会导致鱼类代谢紊乱,饲料蛋白质效率下降,还可引起其他并发症;但饲料脂肪含量过高则会导致脂肪沉积过多,降低抗病力,影响饲料加工。因此饲料中脂肪含量必须适宜。鱼类对脂肪的需要量受食性、生长阶段、饲料中糖类和蛋白质含量及环境因素的影响(麦康森,2017)。
彭墨(2014)以鱼油/豆油(1∶1)为主要脂肪源,配制脂肪含量为3.41%~22.68%的6 种等氮(粗蛋白为51%)等无氮溢出物(13.05%)饲料以探究饲料脂肪水平对大菱鲆[(9.49±0.03)g]生长、脂肪酸组成、血浆生化指标、脂肪沉积及相关基因表达的影响。结果表明,在饲料粗蛋白为50%干重时,脂肪水平在9.378%~15.73%干重均具有较好的生长、饲料利用和生理状态,脂肪水平过高或过低均会降低以上指标;过低或过高饲料脂肪水平均一定程度上促进了肝脏脂肪分解相关基因表达;高脂造成大菱鲆肝脏脂肪沉积可能与脂肪合成上调有关。Zhang等(2015)采用了类似彭墨的试验方法,研究了日粮(粗蛋白为45%)中不同脂肪水平(6.0%~13.5%)对大菱鲆[(39±0.2)g]生长性能、体成分和抗氧化状态的影响。结果表明,高水平的日粮脂肪增加了大菱鲆的生长速度,但生长的增加主要是由于脂肪沉积过多。含13.5%脂肪的日粮引发了大菱鲆的氧化应激。基于大菱鲆的生长和抗氧化性能考虑,大菱鲆日粮脂肪水平以11%为宜。马爱军等(2005)的研究表明,与投喂低蛋白和低水平n-3 HUFA 饲料组相比, 投喂高蛋白和高水平n-3 HUFA 饲料的大菱鲆亲鱼[(2650±12.00)g] 繁殖力明显提高。且经过30 天的饲育,大菱鲆苗种的存活率明显高于低蛋白和低脂肪酸组。且该研究推荐大菱鲆亲鱼培育饲料中脂类的适宜含量≥10%。Leknes 等(2012)探讨了日粮三种粗蛋白水平(35%、42.5%、50%)和两种粗脂肪水平(15%和25%)对大菱鲆(580 g)生长性能和鱼肉品质的影响。尽管大菱鲆(>500 g)的商业饲料中的CP 范围通常为52%~54%,但该研究表明,大菱鲆能够有效地利用高脂饲料,低水平和中等水平的蛋白饲料对大菱鲆具有一定的保蛋白作用。饲料中支持最佳生长的最低CP 和CL 含量分别为43.5%和25.7%。
此外,毕铮铮等(2019)研究了不同脂肪水平(10%、14%、18%)下添加0.09%的胆汁酸对大菱鲆[(23.12±0.10)g]生长和肌肉脂肪酸组成的影响。结果表明,胆汁酸水平为0.09%、饲料脂肪水平为18%时,更能促进大菱鲆生长发育,但不会改变其肌肉脂肪酸组成。孙建珍等(2014)也研究了两个脂肪水平(10%、18%)下,分别添加0%和1.5%的胆汁酸对大菱鲆幼鱼[(45.78±0.11)g]生长、体组成和脂肪代谢的影响。结果表明,在2 个脂肪水平下添加胆汁酸均能促进大菱鲆幼鱼的生长,提高饲料利用,促进脂肪消化吸收,降低组织中粗脂肪含量。唐卓懿等(2017)探究了饲料适宜脂肪含量(11.69%) 和高脂肪含量(16.58%)对大菱鲆幼鱼[(8.6±0.01)g]雷帕霉素受体(TOR)信号通路的影响。结果表明,饲料脂肪水平升高,肝脏TOR 信号通路被激活,肌肉TOR 信号通路受到部分抑制。肝脏和肌肉TOR信号通路的变化规律不同,可能与肌肉对胰岛素的敏感性发生变化有关。
综上,大菱鲆饲料适宜脂肪范围为:幼鱼9.4%~18.0%,中成鱼25.7%,亲鱼≥10%(表1)。

表1 大菱鲆的脂肪需求
1.2 脂肪酸
由于多数脂质为混合物,脂肪酸通常是动物摄食饲料脂质的主要来源(麦康森,2015)。其中必需脂肪酸是指鱼类生长所必需但其本身不能合成,或合成量不能满足需要,必需由饲料直接提供的脂肪酸(麦康森,2017)。张圆琴等(2017)研究了饲料中不同水平(0.72%、5.63%、15.03%)的花生四烯酸(ARA)对发育前期的大菱鲆亲鱼性类固醇激素合成量及合成过程的影响,结果发现,饲料中添加一定量的ARA抑制了发育前期大菱鲆亲鱼雌二醇和睾酮的合成,在卵巢中,这种抑制作用可能是通过抑制促卵泡激素受体的表达来实现,而在精巢中,可能是通过抑制固醇合成急性调节蛋白以及17α羟化酶来实现。综合分析,在大菱鲆性腺发育的前期,建议饲料中ARA 的含量宜低于总脂肪酸的6%。马爱军等(2005)的研究表明,与投喂低蛋白和低水平n-3 高不饱和脂肪酸(HUFA)饲料相比,投喂高蛋白和高水平n-3 HUFA 的饲料,大菱鲆亲鱼[(2650±12.00)g] 繁殖力明显提高,并建议大菱鲆饲料中脂类的适宜含量应≥10%,其中总n-3 HUFA 的适宜比例≥20%。谭青(2017)研究了饲料中6 种不同n-3/n-6 HUFA 比 值(29.54、23.04、18.97、9.06、6.86、3.87)对大菱鲆幼鱼[(12.18±0.01)g]生长性能及饲料利用、体组成和消化酶的影响。综合各因素分析,分别以生长性能和免疫性能为考量指标,得出饲料中n-3/n-6 HUFA 比值的适宜范围为18.97~23.04 和9.06~23.04;且鱼体中ARA 的含量随着饲料中ARA 的增加而增加。C18∶1n-9不能作为大菱鲆EFA(必需脂肪酸)缺乏的指示物。鱼体优先富集二十二碳六烯酸(DHA)、多不饱和脂肪酸(PUFA),ARA 比二十碳五烯酸(EPA)更能富集在鱼体中。谭青等(2018)同时探讨了饲料中不同n-3/n-6 LC-PUFA对大菱鲆幼鱼生长性能及饲料利用、体组成和消化酶的影响,研究表明,饲料中n-3/n-6 LC-PUFA 比值在适宜范围(18.97~23.04)能促进大菱鲆幼鱼的生长,同时,饲料中ARA 对消化呈先上升后降低的趋势,n-3/n-6 LC-PUFA 的比例过低会对大菱鲆幼鱼存在降脂作用。彭墨(2015)的研究则表明,大菱鲆幼鱼[(5.89±0.02)g] 饲料中n-3 LC-PUFA 含量应大于0.8%。此外,大菱鲆全鱼及各部位脂肪酸组成见表2,以供研究者参考。
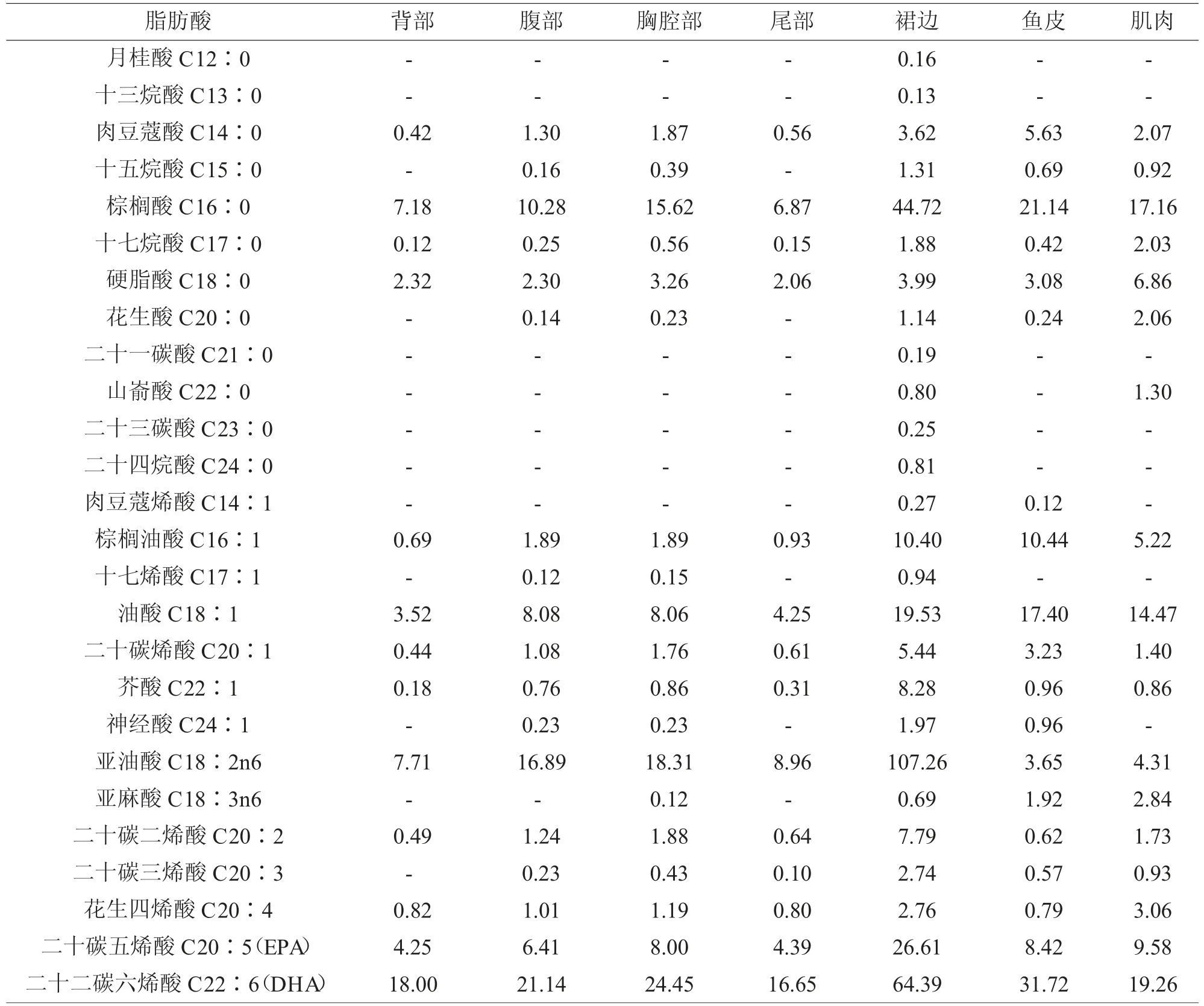
表2 大菱鲆全鱼及各部位脂肪酸组成
1.3 类脂质
1.3.1 磷脂
在饲料中添加完整的磷脂可促进仔稚鱼和早期幼鱼的生长,也能够提高仔稚鱼的成活率并降低畸形率(Tocher 等,2008)。仔稚鱼体内磷脂的合成是受限制的,必须在饲料中添加完整的磷脂,以便于脂肪酸和脂类能够有效地从肠道转运到其他组织(麦康森等,2015)。
Li 等(2018)探讨了不同水平(0~0.55%) 的溶血卵磷脂(LPC)对大菱鲆幼鱼[(41.3±1.6)g]生长、养分表观消化率和脂质代谢的影响。结果表明,日粮LPC 显著提高了生长性能和胆体比,显著降低了脏体比。日粮LPC 降低了能量的表观消化率,但对蛋白质和脂肪没有影响。日粮LPC 提高了脂类代谢相关酶的活性。总之,日粮LPC 降低了大菱鲆的脂肪需要量,提高了大菱鲆的脂肪利用率,综合分析,大菱鲆幼鱼适宜日粮LPC 为870.37mg/kg。刘兴旺等(2012)以鱼粉和大豆浓缩蛋白作蛋白源,配制5 种等氮等能的饲料。5 种饲料分别含有0%、0.5%、1.0%、2.0%和3.0%的大豆卵磷脂以替代相同水平的油脂(鱼油∶豆油为1∶1)。结果发现,饲料中不同水平大豆卵磷脂对大菱鲆[(15.74±0.48)g]的成活率、特定生长率、饲料效率和血清甘油三酯均无显著影响,但随着饲料中大豆磷脂水平的升高,特定生长率和饲料效率有升高的趋势、血清甘油三酯有下降的趋势。同时,随着饲料中大豆磷脂水平的升高,血清总胆固醇呈显著下降趋势。
1.3.2 胆固醇
胆固醇是动物细胞膜的主要结构性成分之一,也是生物体组织中多种功能性物质的前体。Yun 等(2011)和贠彪(2012)研究了高植物蛋白日粮(含14.5%鱼粉、42.0%豆粕和18.5%麦麸粉) 中添加不同水平胆固醇(0.0%、0.5%、1.0%、1.5%)对大菱鲆幼鱼[(5.84±0.02)g] 生长性能、采食量和胆固醇代谢的影响,对照组日粮含58%的鱼粉。对应的处理组日粮胆固醇浓度分别为0.30%、0.77%、1.25%和1.78%,对照组为0.63%。结果表明,高植物蛋白日粮中1.25%的胆固醇水平可显著改善大菱鲆幼鱼的生长性能,且无不利影响。此外,上述高植物蛋白日粮中添加1.0%的胆固醇和1.0%的牛磺酸可显著改善大菱鲆幼鱼的生长性能,并且不产生负面影响(Yun 等,2012;贠彪,2012)。贠彪(2012)的研究同时表明,与摄食全鱼粉组相比(58%的鱼粉),摄食高植物蛋白饲料显著上调了大菱鲆肝脏3- 羟基-3-甲基戊二酸单酰辅酶A 还原酶(HMG-CoAr)基因mRNA 的表达。然而,当高植物蛋白饲料中补充1.0%的胆固醇则显著抑制肝脏基因的表达量。因此,充足的饲料胆固醇能够显著抑制HMG-CoAr 基因mRNA 的表达,胆固醇的合成受反馈调节机制控制。Zhu 等(2014)和朱腾飞(2014)也研究了日粮中添加不同水平(0.0%、0.5%、1.0%、1.5%、2.0%) 胆固醇对大菱鲆幼鱼[(5.18±0.01)g]采食量、生长性能和胆固醇代谢的影响。对应的日粮总胆固醇浓度分别为0.38% 、0.76% 、1.30% 、1.80% 、2.22%。结果表明,饲料中胆固醇含量在1.30%以上时可能对鱼体健康有害。日粮中添加适量的胆固醇可促进大菱鲆的生长发育,但对大菱鲆幼鱼采食量的促进作用有限。
此外,Zhu 等(2014)和朱腾飞(2014)研究了饲料中磷脂和胆固醇的交互作用对大菱鲆幼鱼[(5.18±0.01)g]摄食、生长和胆固醇代谢的影响。研究以三种水平的磷脂(0%、2%、4%)和三种水平的胆固醇(0%、1%、2%)交互制成九组等氮等脂的实验饲料。结果发现,磷脂和胆固醇对大菱鲆幼鱼的增重率具有显著影响,但是对于摄食率的影响并不显著。在胆固醇代谢过程中,饲料磷脂和胆固醇仅在胆固醇运输方面的高密度脂蛋白胆固醇和低密度脂蛋白胆固醇产生了交互作用,对胆固醇合成关键酶HMG-CoA 还原酶和胆固醇降解为胆汁酸过程中关键酶CYP7A1 都没有显著的交互作用,表明了饲料磷脂在胆固醇运输过程中具有重要作用。
1.4 脂肪源替代
水产养殖业可持续发展与鱼油资源有限性的矛盾日益激化,寻找合适的替代脂肪源势在必行。目前,大菱鲆饲料脂肪源替代的研究主要集中于棕榈油(Palm oil)、菜籽油(RO)、豆油(SO)、亚麻籽油(LO)、花生油(PO)和猪油(Lard)。
Wang 等(2016)和王庆超(2015)研究了LO 替代四个水平(0、33.3%、66.7%和100%)的鱼油(FO)对大菱鲆[(5.00±0.10)g]的影响。结果表明,LO 替代66.7%的FO,不影响大菱鲆生长性能、免疫性能和饲料效率。LO完全取代FO,抑制了脂肪酸的β 氧化,促进了脂质合成;但显著降低了血清溶菌酶活性和总抗氧化能力,对超氧化物歧化酶活性则无显著影响。LO 完全取代FO 导致大菱鲆出现轻度贫血。此外,该研究表明,在以亚麻油为基础脂肪源的饲料中添加一种海洋微藻——裂殖壶藻(Schizochytrium sp.),能够显著改善大菱鲆的生长表现以及免疫功能。李思萌等(2015)配制的七种等氮等脂的饲料中分别以6%的FO、SO、RO、PO、Lard、FO-SO (1∶1)(FSO)和FO-SO-Lard(3∶4∶3)的混合油(MIX)作为脂肪源,40 天的养殖试验结果发现,综合以上7 种饲料脂肪源对大菱鲆[(27.31±0.10)g]生长性能、体组成及血清生化指标的影响分析,FSO、SO 和RO 可作为大菱鲆饲料合适的脂肪源。赵曼茜等(2019)等采用类似于李思萌等的方法,研究了饲料脂肪源对大菱鲆幼鱼[(27.31±0.10)g]生长性能和肌肉脂肪酸组成的影响。结果也发现,SO 和RO 是大菱鲆幼鱼饲料良好的脂肪源,FSO 则能使大菱鲆幼鱼更好地生长,而Lard、MIX 不适宜作为大菱鲆的饲料脂肪源。彭墨(2014)和Peng 等(2017)的研究表明,在满足n-3 LC-PUFA 需求下,Palm oil、RO、SO 和LO 替代66.7%FO 并未显著影响大菱鲆[(9.49±0.03)g]生长和饲料利用;饲料相对高MUFA(RO)水平将更有利于大菱鲆的心血管健康;摄食RO、SO 和LO 饲料的大菱鲆肝脏脂肪过多沉积可能是通过上调脂肪合成基因表达。此外,分别以Palm oil、RO、SO、LO分别替代四个水平(0%、33.3%、66.7%和100%)的鱼油,配制13种等氮等脂饲料以探究植物油替代不同水平的鱼油对大菱鲆幼鱼[(5.89±0.02)g]生长、脂肪酸组成以及肝脏和肌肉脂肪沉积的影响。92d 天的养殖试验结果发现,100%SO 日粮组大菱鲆生长性能明显低于低水平SO 日粮组。大菱鲆日粮SO 水平越高,肝脏脂质沉积增加,可能与脂肪酸合成相关基因(FAS)的表达上调以及脂肪酸氧化基因(CPT I)的表达下调有关(Peng 等,2014)。Palm oil、RO、SO 和LO 替代66.7%鱼油均未显著影响大菱鲆生长和饲料利用,但大菱鲆肝脏脂肪含量显著升高,且肌肉中EPA 和DHA 含量显著下降,故从营养品质角度建议大菱鲆饲料中植物油水平应低于66.7%,且大菱鲆饲料中n-3 LC-PUFA含量应大于0.8%(彭墨,2014;彭墨等,2014;彭墨等,2015)。
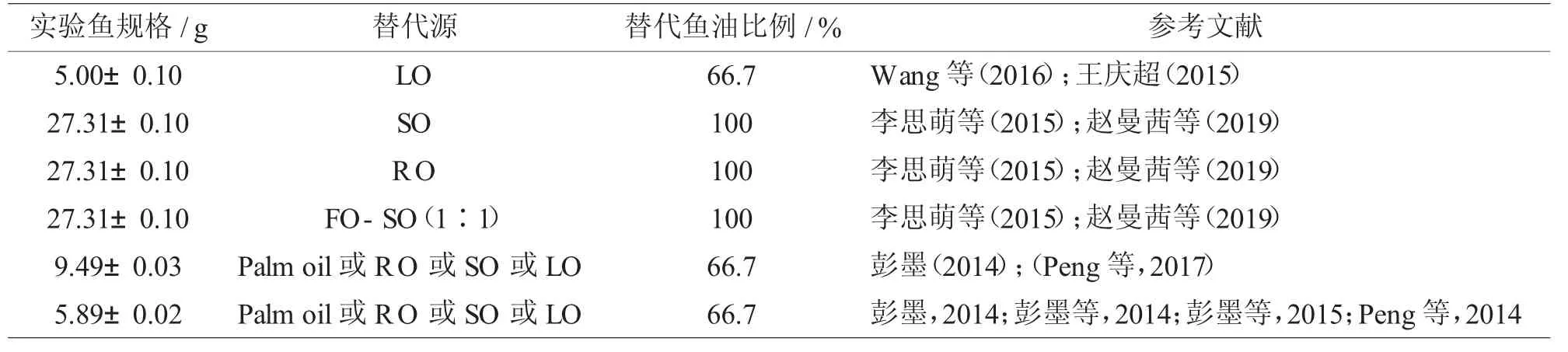
表3 大菱鲆配合饲料中的鱼油替代研究
2 大菱鲆糖类营养
糖类是动物体细胞的组成成分,可为动物提供能量以及合成体脂肪和非必须氨基酸,并具有蛋白质节约效应。虽然相比于蛋白质代谢和脂肪代谢,鱼类(尤其是肉食性鱼类)糖代谢的作用是次要的,但糖类对于所有鱼类而言又是至关重要和无法替代的(麦康森等,2015;Hemre等,2002)。
李晓宁(2011)以鱼粉为蛋白源,鱼油和豆油为主要脂肪源,小麦粉为主要糖源,研究不同饲料糖水平(15%~25.5%)对不同生长阶段(4.30±0.01g、89.57±0.32g、208.30±0.29g)大菱鲆生长、生理状态和体成分的影响,并探究了不同生长阶段大菱鲆饲料中糖的适宜含量。结果表明,随着大菱鲆的生长,其对饲料糖的耐受能力越来越强,大菱鲆幼鱼饲料中的糖的适宜含量为15.0%或更低,而中鱼和大鱼阶段饲料中的糖含量可达25.5%。聂琴等(2013)从饲料糖类对大菱鲆[(8.06±0.08)g]代谢途径影响的角度,分析了其糖代谢酶活性和相对表达量的变化情况。结果表明,饲料中添加15%的葡萄糖或糊精时,可以有效促进大菱鲆肝脏糖酵解能力。蔗糖作为饲料糖源时,仅在添加量为28%时显著促进糖酵解酶GK 活性,糖酵解其他酶活性以及糖异生酶活性均不受蔗糖水平的显著影响。此外,糊精对大菱鲆肝脏糖异生酶的基因表达存在一定程度的抑制,但对葡萄糖-6- 磷酸酶的两种亚型都无影响;添加葡萄糖或蔗糖都对上述酶的基因表达无影响;大菱鲆肝脏糖异生关键酶磷酸烯醇式丙酮酸羧激酶(线粒体型)的表达不受饲料糖的影响,为组成型表达;大菱鲆中糖代谢酶的关键调控位点在进化过程中均保守存在。苗淑彦等(2013)和Miao等(2016)评估了在低蛋白水平(40%)下,饲料中不同种类的碳水化合物[葡萄糖(15%)、蔗糖(15%)和糊精(15%)]对大菱鲆幼 鱼 [(8.12±0.04)g、(8.06±0.08)g]生长、成活、饲料利用、体组成和血液生理生化指标等的影响。综合生长性能、三种碳水化合物的表观消化率以及糖代谢分析得出,大菱鲆对糊精的利用效率高于葡萄糖和蔗糖。并且不同种类的碳水化合物通过糖脂关联代谢等途径对大菱鲆幼鱼的体组成和血液生理生化指标造成了一定的影响。苗慧君(2013)和苗慧君等(2014)在饲料中添加不同水平的糊精(0%、5%、15%),分别替代不同水平(50%、45%、40%)的蛋白质以研究糖替代蛋白对大菱鲆肝脏中脂肪合成相关酶活性、脂肪分解相关酶活性以及肝脏中脂肪酸组成的影响。结果表明,15%的糊精添加水平显著升高了葡萄糖-6- 磷酸脱氢酶和肠道脂肪酶的活性;此外,饲料中糖替代蛋白对脂蛋白脂酶、肝脂酶、总脂酶的活性以及肌肉中脂肪酸组成的影响不大,对单不饱和脂肪酸中C16∶1 有显著影响,对多不饱和脂肪酸、高不饱和脂肪酸等均没有显著影响。

表4 大菱鲆的部分营养需要
3 大菱鲆日粮中蛋白质、脂质与碳水化合物之间的关系
组成蛋白质的各种氨基酸均可在体内转化成脂肪;脂肪可转化为糖类,但脂肪酸不能合成糖类;蛋白质也可在鱼体内转化成糖类;这三大营养物质均可在体内氧化供能(麦康森等,2015)。
Liu 等(2014)评估了三种蛋白质水平(45、50 和55%)、两种脂质水平(12.0 和16.0%)、P:E范 围 为87.4~110.0mg 蛋 白 质/kcal 的六种实际试验日粮,对大菱鲆在3 个不同生长阶段(4.5±0.01g、59.1 ±0.24g 和 209.1 ±0.21g)的生长、饲料利用率和体成分的影响,试验时间9 周。结果表明,无论日粮脂肪是何水平,随着日粮蛋白质水平的增加,大菱鲆生长性能普遍提高。在同一蛋白质水平下,随着脂肪含量的增加,饲料效率比和蛋白质效率比均显著增加,大菱鲆大鱼组[(209.1±0.21)g]除外。大菱鲆中成鱼[(59.1±0.24)g]的特定生长率也随着脂肪水平的增加而提高,这表明脂肪具有蛋白质节约效应。Miao 等(2016)配制了糖脂比分别为2∶18、6∶18、18∶12 和28∶6 的四种等氮等能的低蛋白(39%)日粮,蛋白水平50%,糖脂比为2∶12 的日粮为对照。研究日粮碳水化合物与脂质比例(CHO∶LIP)对大菱鲆幼鱼[(8.75 ±0.04)g]生长性能和饲料利用的影响,试验为期9周。实验结果基于生长性能考虑,日粮蛋白质水平为39%时,适宜的CHO∶LIP 比例为18∶12。Zeng 等(2015)研究了五种盐度(12、18、24、30、36)下大菱鲆幼鱼日粮中碳水化合物对蛋白质的保护作用。试验配制了三种等热量和等脂的饲料,为期60天。结果表明,在盐度为18~30时,将大菱鲆日粮中糊化淀粉含量从5%增加到15%可以节约6%的日粮蛋白质,而较高的盐度(36)可以提高日粮碳水化合物的利用率,增强蛋白质节约效应,这可能是因为盐度为36 时,日粮淀粉可诱导脂肪生成。
瘦素是一种脂肪细胞分泌的蛋白类激素,脂联素是主要由脂肪组织分泌的一种胶原样脂肪因子,两者均能抑制肝脏中的糖异生过程。韩冬冬(2015)和韩冬冬等(2017)以糊精为主要糖源,鱼油和大豆卵凝脂为主要脂肪源,白鱼粉、酪蛋白和明胶为蛋白源,研究了不同饲料糖脂比对大菱鲆[(8.06±0.08)g]瘦素及其受体和脂联素受体基因表达的影响。结果表明,饲料糖脂比从1∶6 升高到2∶1 对瘦素和瘦素受体基因表达没有产生显著影响,但在糖脂比上升到14∶1 时,肝脏瘦素表达量显著升高,而肝脏瘦素受体的mRNA 水平却显著低于对照组。饲料糖脂比的变化可能通过改变脂联素受体的表达量,从而间接影响大菱鲆体内脂联素的作用。
4 总结与展望
综合大菱鲆脂肪及糖类需求的相关研究成果,结合高效安全环保型大菱鲆配合饲料的要求,大菱鲆各营养素推荐添加范围见表4。此外,相关研究表明,棕榈油、菜籽油、豆油、亚麻籽油单独或混合替代大菱鲆配合饲料中66.7%的鱼油是可行的。
目前,大菱鲆脂类及糖类的营养需求与饲料研究仍然存在较多不足,例如有关大菱鲆对脂肪及糖类的需求研究主要集中于幼鱼,中鱼与大鱼的相关研究则较少。配合饲料中脂肪酸的具体可用数据较少,在不同生长阶段和不同养殖模式下的脂肪酸需求参数和特点尚不清晰。为降低水产养殖过程中对鱼油的依赖,需深入研究其他动植物脂肪源对鱼油的替代,以促进大菱鲆养殖业的健康可持续发展。
参考文献:(略)
