乳酸片球菌AS185调节高脂高糖饮食诱导的代谢综合征
2021-01-20赵子健段翠翠赵玉娟李盛钰
张 麟,高 磊,王 超,赵子健,段翠翠,赵玉娟,杨 舸,李盛钰,*
(1.吉林省农业科学院农产品加工研究所,吉林 长春 130033;2.吉林农业大学食品科学与工程学院,吉林 长春 130118)
代谢综合征是一组复杂的代谢紊乱症候群,是城市化和生活方式的改变,特别是将常规饮食改为高脂、高糖饮食造成的[1]。代谢综合征是导致肥胖、高血脂、糖尿病和心血管疾病等的危险因素[2],因此其诊断和治疗方案也越来越受到重视。
益生菌被定义为公认食用安全的活的微生物,摄入足够数量时,对宿主的健康有促进作用。益生菌是一类重要的功能性食品原料,因其在调节肠道菌群、改善便秘和腹泻、抑制有害微生物、调节免疫系统功能、预防肿瘤等方面的潜在功效而受到消费者的关注[3-7]。特别是其在减肥、调节血脂和降低血清胆固醇、降血糖、抗动脉粥样硬化等代谢调控紊乱相关疾病防治方面的作用[8-12]。
乳酸片球菌(Pediococcus acidilactici)是一种革兰氏阳性球菌,在商业发酵过程中广泛用于生产普通食品[13]。有研究报道,通过口服乳酸片球菌R037可以减轻慢性炎症并调节免疫反应,成功缓解了动脉粥样硬化[14-15]。乳酸片球菌AS185是本实验室自主分离鉴定的一株乳酸菌新菌株,前期研究表明,该菌株具有较好的黏附和耐受能力,并通过调节血脂、抑制炎症因子及黏附分子改善抗动脉粥样硬化症状[16]。因此,推测乳酸片球菌AS185具有减轻不健康的血脂异常状态和抑制炎症的能力,可能适用于改善代谢综合征,但其作用和机制还需进一步深入研究。本研究考察乳酸片球菌AS185对高脂高糖诱导代谢综合征模型小鼠的治疗作用和机制,为其开发成食品、保健品、药品等提供理论依据。
1 材料与方法
1.1 动物、菌株与试剂
45 只7 周龄(体质量35~50 g)清洁级ICR雄性健康小鼠,购于长春市亿斯实验动物技术有限公司,生产许可证号:SCXK(吉)2011-0004。进行为期1 周的适应性喂养,动物房温度(21±2)℃、相对湿度(40±10)%,采食和饮水由小鼠自由进行。
基础饲料和高脂饲料(83.3%(质量分数,下同)基础饲料+10%猪油+3%胆固醇+3%白砂糖+0.5%的胆酸钠+0.2%丙基硫氧嘧啶)均由长春市亿斯实验动物技术有限责任公司提供。
实验菌株AS185是从吉林省延吉地区传统农家大酱中分离获得的一株乳酸菌新菌株,经API 50 CHL鉴定系统和16S rDNA序列比对,鉴定为乳酸片球菌(Pediococcus acidilactici)。菌株保藏于中国典型培养物保藏中心,保藏登记号为CCTCCNO:M2018114[17]。
总胆固醇(total cholesterol,TC)、甘油三酯(triglyceride,TG)、高密度脂蛋白胆固醇(highdensity lipoprotein cholesterol,HDL-C)、低密度脂蛋白胆固醇(low-density lipoprotein cholesterol,LDL-C)检测试剂盒 南京建成生物技术公司;小鼠肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素-1β(interleukin-1β,IL-1β)、白细胞介素-6(interleukin-6,IL-6)、胰岛素(insulin,INS)、游离脂肪酸(free fatty acids,FFA)、C-反应蛋白(C-reactive protein,CRP)、脂多糖(lipopolysaccharide,LPS)酶联免疫吸附检测试剂盒 上海江莱生物科技有限公司;二喹啉甲酸(bicinchoninic acid,BCA)蛋白定量试剂盒北京鼎国昌盛生物技术有限公司;兔抗核因子κ B(nuclear factor kappa-B,NF-κB)抗体、兔抗磷酸化NF-κB(phosphorylation NF-κB,p-NF-κB)、兔抗Toll样受体4(Toll-like receptor 4,TLR4)单克隆抗体、兔抗髓样分化因子(myeloid differentiation factor 88,MyD88)单克隆抗体、兔抗肝激酶B1(liver kinase B1,LKB1)单克隆抗体 英国Abcam公司;腺苷酸活化蛋白激酶(adenylate activated protein kinase,AMPK)、磷酸化AMPK(phosphorylation AMPK,p-AMPK)、乙酰辅酶A羧化酶(acetyl-CoA carboxylase,ACC)、磷酸化ACC(phosphorylation ACC,p-ACC)、固醇调节元件结合蛋白1c(sterol regulatory element-binding protein-1c,SREBP-1c)单克隆抗体 北京博奥森生物技术有限公司;山羊抗兔二抗 北京中杉金桥生物技术有限公司。
1.2 仪器与设备
超高速冷冻离心机 美国Thermo公司;血糖仪瑞士Roche有限公司;ELx800全自动酶标仪 美国BioTek公司;DHP-9272型电热恒温培养箱 上海一恒科技有限公司;SQ810C型高温高压灭菌锅 重庆雅玛拓科技有限公司。
1.3 方法
1.3.1 菌悬液的制备
连续活化3 代的乳酸片球菌AS185菌株接种于MRS液体培养基中,经37 ℃静置培养16 h后,将菌液在4 ℃、4 000×g条件下离心15 min,弃上清液,取菌泥用灭菌生理盐水洗涤1 次,并调整活菌数浓度为1×109CFU/mL,备用。
1.3.2 造模方法与分组
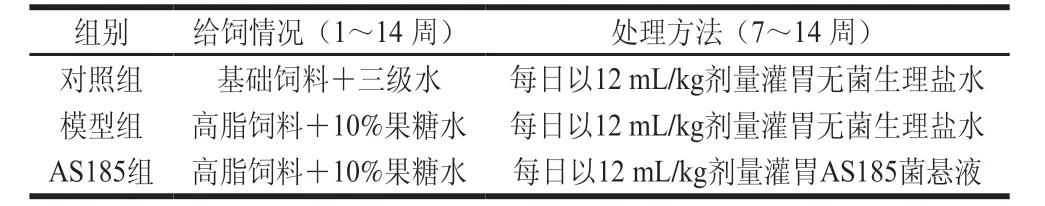
表1 动物实验方案Table 1 Grouping of experimental animals
按参考文献[18]的方法,小鼠随机分为3 组,每组15 只(表1)。每2 周测体质量并尾静脉取血测定断食不断水12 h的空腹血糖(fasting blood glucose,FBG)浓度1 次。造模结束后,禁食12 h后,尾静脉取血测FBG测定、血脂水平(TC、TG、LDL-C、HDL-C)并检测体质量。饲养6 周结束后,模型组和AS185组小鼠满足以下条件中任意2 项即为代谢综合征模型造模成功小鼠[19]:1)体质量高于对照组小鼠体质量20%;2)血清中血脂水平改变,即TC、TG、LDL-C水平升高和HDL-C水平降低;3)胰岛素抵抗指数(insulin resistance index,IR)升高。最终,各组筛选出10 只符合模型纳入标准的小鼠用于后续实验。其中,AS185组灌胃乳酸片球菌AS185菌悬液(1.0×109CFU/mL),剂量为每日12 mL/kg,对照组和模型组给予等体积的无菌生理盐水灌胃,连续灌胃8 周(第7~14周)。每2 周测定体质量及FBG浓度。末次灌胃后,眼球采血,4 ℃、3 000×g条件下离心10 min,收集血清,-80 ℃冻存;断颈处死后,解剖,摘取肝脏,-80 ℃冻存,备用。
1.3.3 血清生化指标测定
血脂指标(TC、TG、HDL-C、LDL-C和FFA)、血清炎症指标(TNF-α、IL-6、IL-1β、CRP和LPS)及INS水平检测,按照试剂盒说明书方法操作。利用稳态模型(homeostasis model assessment,HOMA)评估胰岛素抵抗(insulin resistance,IR)指数(HOMA-IR),HOMA-IR按照下式计算。

1.3.4 Western Blot检测蛋白表达
根据文献[20]的方法,取各组小鼠肝脏50 mg,加入适量蛋白裂解液匀浆,离心收集上清液,采用BCA法测定总蛋白质量浓度。首先,加上样缓冲液同时沸水浴10 min使蛋白变性,-20 ℃冷藏,备用;之后进行蛋白电泳分离,12%聚丙烯酰胺凝胶电泳(浓缩胶60 mV电压,30 min;分离胶120 mV电压,120 min),随后60 mV电压,90 min转膜至PVDF膜;室温封闭60 min,3%牛血清白蛋白封闭液于摇床上进行封闭;最后孵育一抗,TBST洗膜3 次,每次7 min,并分别加入TLR4、NF-κB、p-NF-κB、LKB1、AMPK、p-AMPK、ACC、p-ACC、SREBP-1c等抗体,4 ℃反应过夜;次日,以TBST洗膜3 次,每次10 min;再加入HRP标记的山羊抗兔IgG(稀释体积比为1∶3 000),常温下反应1 h;再用TBST洗膜4 次,每次5 min;采用电化学发光法曝光,并用Image J软件进行半定量分析。
1.4 数据统计分析
分析处理数据使用GraphPad Prism 5软件进行绘图,使用SPSS 23.0软件的单因素方差分析对数据进行差异显著性比较,结果均以±s表示。
2 结果与分析
2.1 乳酸片球菌AS185对代谢综合征模型小鼠体质量、INS、FBG及HOMA-IR的影响
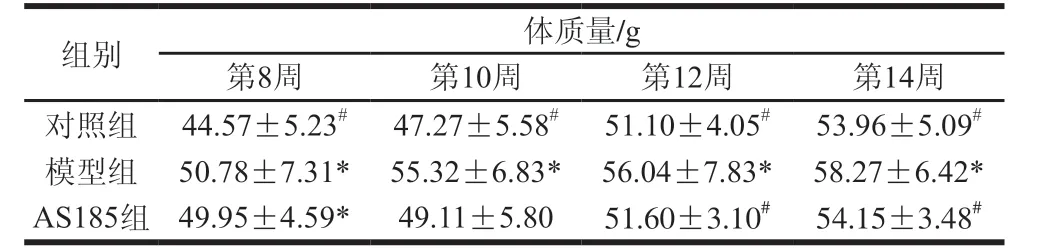
表2 乳酸片球菌AS185对高脂高糖诱导代谢综合征模型小鼠体质量的影响Table 2 Effect of P.acidilactici AS185 on body mass of mice with metabolic syndrome induced by high-fat and high-fructose diet
由表2可知,AS185组体质量虽呈上升趋势,但与模型组相比,上升趋势缓慢,并在第12周和第14周体质量与模型组相比出现显著差异(P<0.05),说明乳酸片球菌AS185具有缓解高脂高糖饮食诱导代谢综合征的体质量上升的趋势。

表3 乳酸片球菌AS185对代谢综合征模型小鼠INS、FBG、HOMA-IR的影响Table 3 Effect of P.acidilactici AS185 on INS, FBG and HOMA-IR of mice with metabolic syndrome induced by high-fat and high-fructose diet
由表3可知,14 周后,与模型组相比,AS185组小鼠血清中的INS和FBG水平也呈下降趋势,同时HOMA-IR降低,差异具有统计学意义(P<0.01),说明乳酸片球菌AS185对代谢综合征具有改善胰岛素抵抗的作用。
2.2 乳酸片球菌AS185对高脂高糖诱导代谢综合征模型小鼠血脂水平的影响
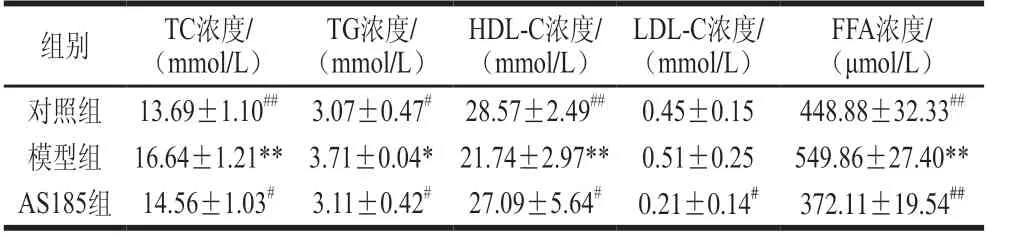
表4 乳酸片球菌AS185对代谢综合征模型小鼠血清TC、TG、LDL、HDL、FFA水平的影响Table 4 Effect of P.acidilactici AS185 on serum TC, TG, LDL, HDL and FFA levels of mice with metabolic syndrome induced by high-fat and high-fructose diet
由表4可知,高脂高糖饮食诱导的代谢综合征小鼠血脂代谢出现紊乱,与对照组比较,模型组小鼠血清中TC、TG、LDL-C及FFA的水平均显著升高,差异具有统计学意义(P<0.05或P<0.01);同时,HDL-C水平有降低趋势;而与模型组相比,AS185组小鼠血脂指标TC、TG、LDL-C、FFA的水平也均降低,差异具有统计学意义(P<0.05或P<0.01),这说明乳酸片球菌AS185能够改善代谢综合征小鼠的血脂代谢紊乱。
2.3 乳酸片球菌AS185对代谢综合征模型小鼠血清炎症因子水平的影响
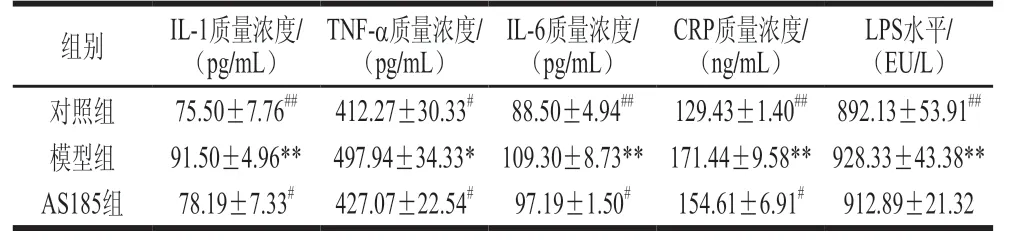
表5 乳酸片球菌AS185对高脂高糖诱导代谢综合征模型小鼠血清炎症因子水平的影响Table 5 Effect of P.acidilactici AS185 on inflammatory factor levels of mice with metabolic syndrome induced by high-fat and high-fructose diet
由表5可以看出,代谢综合征会诱发小鼠体内慢性炎症,与对照组相比,模型组小鼠的血清中炎症因子IL-1β、TNF-α、IL-6、CRP及LPS的水平显著升高(P<0.05或P<0.01);与模型组相比,AS185组小鼠血清中的IL-1β、TNF-α、IL-6、CRP、LPS水平均有降低趋势,其中IL-1β、TNF-α、IL-6、CRP差异具有统计学意义(P<0.05或P<0.01)。以上结果提示,乳酸片球菌AS185有改善代谢综合征小鼠血清中的炎症水平,进而缓解机体慢性炎症状态的可能。
2.4 乳酸片球菌AS185对代谢综合征模型小鼠脏组织蛋白表达的影响
2.4.1 肝组织中TLR4/MyD88/p-NF-κB炎症通路蛋白的表达
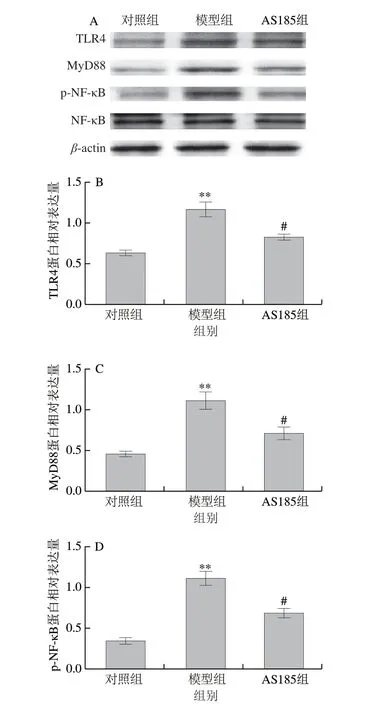
图1 乳酸片球菌AS185对高脂高糖饮食诱导的代谢综合征模型小鼠肝脏炎症通路蛋白表达的影响Fig.1 Effect of P.acidilactici AS185 on the expression of proteins associated with inflammatory signaling pathways in liver of mice with metabolic syndrome induced by high-fat and high-fructose diet
由图1可知,与对照组相比,模型组小鼠肝脏中TLR4、MyD88及p-NF-κB蛋白表达水平极显著上升(P<0.01),表明代谢综合征诱发了肝脏的炎性反应,使机体出现慢性炎症;经乳酸片球菌AS185治疗后,小鼠肝脏中TLR4、MyD88及p-NF-κB表达水平显著降低(P<0.05),NF-κB总蛋白水平无显著变化,说明p-NF-κB水平的改变并不是因为NF-κB总蛋白表达的变化而引起的,而是因为上游MyD88活化引起,说明乳酸片球菌AS185能抑制高脂高糖饮食诱导中TLR4、MyD88及p-NF-κB的表达,降低了肝脏的炎性损伤,缓解机体慢性炎症。
2.4.2 肝组织中LKB1/AMPK/p-ACC脂代谢通路蛋白的表达
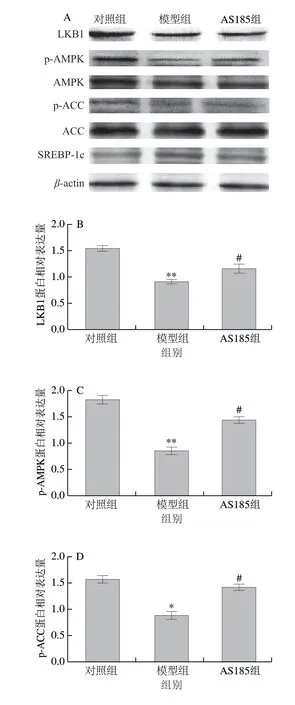

图2 乳酸片球菌AS185对代谢综合征模型小鼠肝组织脂代谢通路蛋白表达的影响Fig.2 Effect of P.acidilactici AS185 on the expression of proteins associated with lipid metabolism signaling pathways in liver of mice with metabolic syndrome induced by high-fat and high-fructose diet
由图2可知,与对照组相比,模型组小鼠肝脏中LKB1、p-AMPK、p-ACC表达水平显著降低(P<0.05或P<0.01),SREBP-1c水平极显著升高(P<0.01),表明高脂高糖能够引起小鼠的脂代谢紊乱;结合图1进一步说明,机体出现慢性炎症的同时会伴随脂代谢紊乱的出现。经乳酸片球菌AS185治疗后,小鼠肝脏中LKB1、p-AMPK和p-ACC表达水平显著升高,SREBP-1c蛋白水平显著降低(P<0.05)。以上结果表明,乳酸片球菌AS185能够干扰LKB1介导高脂高糖对代谢综合征小鼠诱导的肝脏AMPK和ACC磷酸化的作用,并抑制SREBP-1c成熟,改善机体脂代谢的紊乱。
3 讨 论
高糖高脂饮食导致的胰岛素抵抗与系统性低水平炎症密不可分。Mansyur等[21]发现,胰岛素抵抗的发展与炎症有关,几种炎症标记物表现出临床重要性,包括CRP、白细胞、纤维蛋白原和白蛋白,高三酰甘油血症作为代谢综合征成分也会影响炎症。同时也发现胰岛素抵抗与肥胖之间也存在联系,其中脂肪组织在发展胰岛素抵抗的机制中发挥重要作用。脂肪细胞产生的非酯化脂肪酸可通过底物竞争和受损的细胞内胰岛素信号传导来阻止碳水化合物的代谢。脂肪组织分泌的最常见的细胞因子是脂联素,脂联素具有显著的胰岛素增敏作用,因此脂肪组织的增加导致与胰岛素抵抗和代谢综合征相关的脂联素水平的降低。脂肪组织还产生几种炎症因子,例如IL-6,IL-6刺激CRP的产生,这也可以加剧与肥胖有关的炎症,继而导致肥胖相关的胰岛素抵抗[22-23]。与已有研究结果一致,本实验得出高脂高糖饮食的代谢综合征小鼠表现出一系列的代谢紊乱,包括空腹血糖和空腹胰岛素水平均显著增加,血清中TNF-α、IL-1β、IL-6、CRP水平的增加,导致小鼠肥胖出现系统性低水平炎症进而引起血糖-胰岛素稳态失衡,经乳酸片球菌AS185干预后,机体低水平炎症有所改善,同时胰岛素抵抗指数显著下降。
AMPK是哺乳动物细胞能量代谢的重要调节因子。活化的AMPK刺激ATP产生的分解代谢途径,如脂肪酸β氧化,并抑制ATP消耗过程,如脂肪生成[21-22]。AMPK的磷酸化依赖于上游LKB1的激活。LKB1是一种能够磷酸化AMPK的上游激酶,是细胞对低能量反应的关键介质[23]。本研究的结果表明,AS185可能通过LKB1特异性siRNA刺激LKB1磷酸化,并清除LKB1,明显阻断了AS185处理肝细胞的AMPK和ACC磷酸化,通过促进新生脂肪的形成和脂肪酸氧化,可改善高糖高脂饮食诱导的代谢综合征。AMPK的第一个下游酶靶点是ACC,它参与丙二酰辅酶A的合成。AMPK通过磷酸化ACC在Ser77和Ser79上抑制ACC活性,从而刺激脂肪酸氧化和降低脂肪酸合成[24]。此外,乳酸片球菌AS185对SREBP-1c的成熟有抑制作用。SREBP-1c是一种重要的成脂转录因子,在哺乳动物肝脏中含量丰富。由于SREBP-1c的过度表达,脂肪酸进入肝细胞会诱导新的成脂过程[25]。成熟后,SREBP-1c转移到细胞核中,转录上调SCD-1和FAS表达,这是肝细胞从头合成脂肪酸和TG所需的关键酶[26]。本实验发现AS185组TC和FFA的产生减少可能是SREBP-1水平降低所致,这与以往的研究结果[27]一致。因此,本研究结果表明,乳酸片球菌AS185的补充是通过AMPK途径介导ACC磷酸化和抑制SREBP-1信号来达到降脂的作用。
本实验结果表明乳酸片球菌AS185不仅通过调节脂类代谢通路,而且通过在肝水平限制TLR4和MyD88的转录和表达的增加,抑制NF-κB活化,从而减少代谢综合征引起的炎症及其介质。Wang Yuhua等[28]报道,鼠李糖乳杆菌LGG通过抑制TLR4和TLR5以及抑制NF-κB活化和减少TNF-α生成,增加酒精性肝炎症的保护作用。而Jung等[29]的研究表明L.sakeiK040706通过激活巨噬细胞中的TLR2-NF-κB途径产生NO、TNF-α和IL-6,且与TLR4无相互作用。L.paracasei通过诱导NF-κB信号通路的负性调节因子,以TLR2-IRAK4依赖的方式抑制单核细胞顶体产生促炎性细胞因子[30]。另外,Raso等[31]指出L.paracaseiB21060在TLR2和TLR4表达上的活性相同,共享导致NF-κB活化的相同信号级联。因此,益生菌可能通过诱导TLR2、TLR4等多种TLRs受体途径调控高糖高脂饮食诱导的代谢综合征中发挥作用,乳酸片球菌AS185改善代谢综合征引起的炎症反应的准确机制还有待于进一步证实。
综上所述,乳酸片球菌AS185通过MyD88依赖的TLR4/NF-κB通路抑制了促炎症细胞因子的表达,并依赖AMPK通路提升了LKB1、AMPK和ACC蛋白表达的脂代谢通路,改善了高糖高脂饮食诱导的代谢综合征。
