双孢蘑菇培养料木质纤维素利用及相关酶活性变化的研究
2021-01-08杨亚茹张国庆陈青君蔡盼盼王清辉杨世丽
杨亚茹,张国庆,陈青君*,蔡盼盼,王清辉,杨世丽
(1.北京农学院植物科学技术学院/农业应用新技术北京市重点实验室,北京 102206;2.承德兴春和农业股份有限公司,承德 068250;3.邢台市蔬菜站,邢台 054000)
双孢蘑菇(Agaricusbisporus)简称双孢蘑、双孢菇,俗称蘑菇[1]。栽培双孢蘑菇的培养料(堆肥)由作物秸秆(如黍草、稻草和玉米秸秆)、动物粪便(如马粪、牛粪和鸡粪)和石膏等通过隧道发酵而成[2-3]。培养料的理化性质通常作为实际生产中监控其质量优劣、预测产量的重要指标。荷兰是双孢蘑菇栽培技术最为先进的国家,对培养料理化参数有科学的评价指标[4]。中国的双孢蘑菇工厂化生产有了飞速的发展,利用不同的原料制作培养料均获得较好的效果[5]。
双孢蘑菇培养料的主要成分秸秆是木质纤维素,其生产过程是对木质纤维素的降解转化过程[6-7]。实现对木质纤维素的充分降解利用,是工厂化蘑菇生产面临的重要课题[5]。木质纤维素由木质素、纤维素和半纤维素构成,依据其组成元件,可将木质纤维素降解酶划分为纤维素降解酶系、半纤维素降解酶系和木质素降解酶系[8]。纤维素降解酶系包括羧甲基纤维素酶(carboxymethyl cellulase,CMC)、α-纤维二糖酶(α-cellobiase)等;半纤维素降解酶系包括木聚糖酶(xylanase)、木糖苷酶(xylosidase)、α-阿拉伯呋喃糖苷酶(α-arabinofuranosidase)等;木质素的主要降解酶为漆酶(laccase)等;堆肥中酶的活性影响纤维素、半纤维素的降解[9],这些大分子物质必须经相应的酶分解成葡萄糖、果糖等单糖类和小分子化合物后才能被吸收利用,为双孢蘑菇生长提供所需的碳源[10]。以小麦秸秆为主料在一次发酵期间,纤维素和木聚糖被降解程度有限;在二次发酵期间,高达50%的物料碳水化合物被代谢[11]。Chen等[12]、Yama等[13]报道,在堆肥过程中纤维素被降解,在出菇时期主要利用木质素。李晓博等[14]发现酶活性与木质纤维素降解呈正相关,酶活性高降解速率就快,酶活性低降解速率就慢。尽管前人对木质纤维素组分变化及部分酶活性有研究报道,但对于双孢蘑菇全部生产过程中木质纤维素利用规律尚不清晰。
该研究采用隧道式发酵技术,在测定堆肥时期(建堆、一次发酵的3次翻堆、二次发酵)和栽培时期(菌丝长满,原基形成,第1潮菇、第2潮菇、第3潮菇)理化指标(pH值、EC值、含水量、含碳量、含氮量、灰分含量、碳氮比)的基础上,对木质纤维素各组分含量和酶活性进行系统分析,同时统计产量和生物学效率,为双孢蘑菇工厂化栽培建立更为科学的指标评价体系奠定理论基础。
1 材料与方法
1.1 材 料
试验在河北省承德兴春和农业股份有限公司双孢蘑菇工厂进行,双孢蘑菇菌种为A15。
1.2 试验方法
1.2.1 堆肥和双孢蘑菇栽培 采取3批次培养料,试验原料按照碳氮比(30∶1)[15]的原则配方。
第1批次(T1)配方是黍草30 t、稻草30 t、鸡粪60 t、豆粕2 t、石膏4 t;
第2批次(T2)配方是黍草35 t、稻草30 t、鸡粪60 t、豆粕2 t、石膏4 t;
第3批次(T3)配方是黍草36 t、稻草24 t、鸡粪55 t、豆粕2 t、石膏4 t。
通过双孢蘑菇专用空调和PLC智能化控制系统严格控制和调节不同生长时期的温度和湿度、CO2浓度和风压风量。菌丝生长时期培养料温度控制在23~25 ℃,空气达到饱和含水量,CO2保持在0.5%~1%;子实体生长时,菇房室内温度保持在16~18 ℃,空气相对湿度85%~95%,CO2保持在0.08%左右。每批次菇床面积为900 m2,统计每批次菇房产量。
1.2.2 取 样 堆肥时期培养料取样方法:分别在建堆(BM)、一次发酵(PI)(包括2次倒仓PI-1、PI-2和一次发酵结束PI-3)和二次发酵结束后(PII)取样。取样时在隧道前中后的断面上分8个点共采集培养料1~2 kg,培养料混合均匀后部分用于基本理化性质测定,约100 g保存在液氮中,用于相关降解酶活性测定。部分培养料置于-20 ℃冰箱保存,用于培养料木质纤维素组分含量测定。
栽培时期培养料取样方法:在菌丝长满(Filling),原基形成(Pinning),第1潮菇、第2潮菇、第3潮菇(1st、2nd、3rd)取样,取样点在第2层床架的随机3个位点,采样深度为培养料上中层10 cm(除去覆土层),取样300 g左右,将3点的培养料混合均匀。取部分培养料用于基本理化性质测定,约100 g保存在液氮中,用于相关降解酶活性测定。余下培养料置于-20 ℃冰箱保存,用于培养料木质纤维素组分含量测定。
1.2.3 培养料理化性质的测定 使用pH计和电导率仪测定pH值和EC值。采用干燥称重法测定含水量、半自动凯氏定氮法测定含氮量,灼烧重量法测定含碳量和灰分并计算碳氮比。
1.2.4 木质纤维素含量测定 采用滤袋法测定培养料的纤维素和半纤维素含量,Jurak[16]的方法测定木质素的含量。
1.2.5 木质纤维素相关酶活性的测定 粗酶液的制备:取3.0 g培养料,加入30 mL的生理盐水,25 ℃、220 r/min振荡悬摇2 h,4层纱布过滤后,4 ℃、12 000 r/min离心10 min,收集上清液。
以羧甲基纤维素钠作为降解底物测定粗酶液的羧甲基纤维素酶活性;以木聚糖作为降解底物测定粗酶液的木聚糖酶活性,酶活性的定义以每分钟释放1 μmol的葡糖糖/木糖所需要的酶量(U/g);以对硝基苯酚为降解底物,测定α-纤维二糖酶、木糖苷酶、α-阿拉伯呋喃糖苷酶、木聚糖酶的活性,酶活性的定义以每分钟释放1 μmol的对硝基苯酚所需要的酶量(U/g)。运用ABTS法测定粗酶液的漆酶活性,酶活性的定义以每分钟、每毫升反应体系产生一个吸光值所需要的酶量(U/g)。
1.2.6 计算双孢蘑菇产量和生物学效率 分别统计每个生产批次每潮菇的产量,并计算双孢蘑菇的生物学效率。
生物学效率(%)=蘑菇鲜质量(g)/培养料干质量(g)×100%
2 结果与分析
2.1 培养料的理化性质
保持在适宜范围的理化参数是双孢蘑菇正常产出的前提。表1为双孢蘑菇工厂化生产时使用3批次基本相同的配方在堆肥时期的理化性质。pH值在一次发酵时先升高后降低,尤其是第1批次和第2批次都是从堆肥时期就开始升高到第2次翻堆pH达到最高分别是9.42和8.03,然后从一次发酵结束(PI-3)到二次发酵(PII)结束后pH逐渐降低。EC值从建堆(BM)到一次发酵(PI)再到二次发酵(PII)都呈逐渐下降趋势,堆肥时期EC值变化不大;含水量从建堆(BM)到一次发酵(PI)保持在73.53%~77.03%,经过二次发酵结束之后(PII)降至67.93%;含碳量整体呈下降趋势,从建堆(BM)到一次发酵第1次翻堆(PI-1)稍有升高,之后逐渐下降;含氮量从建堆(BM)到二次发酵(BM~PII)逐渐增加,从1.37增加到1.98;灰分含量整体趋势逐渐增加,但是从建堆(BM)到一次发酵第1次翻堆(PI-1)稍有降低,之后逐渐升高。碳氮比从建堆的29.21到二次发酵结束的17.7呈下降趋势。3批次培养料理化性状指标均在适宜范围,为产量形成和木质纤维素分析提供良好的基础。
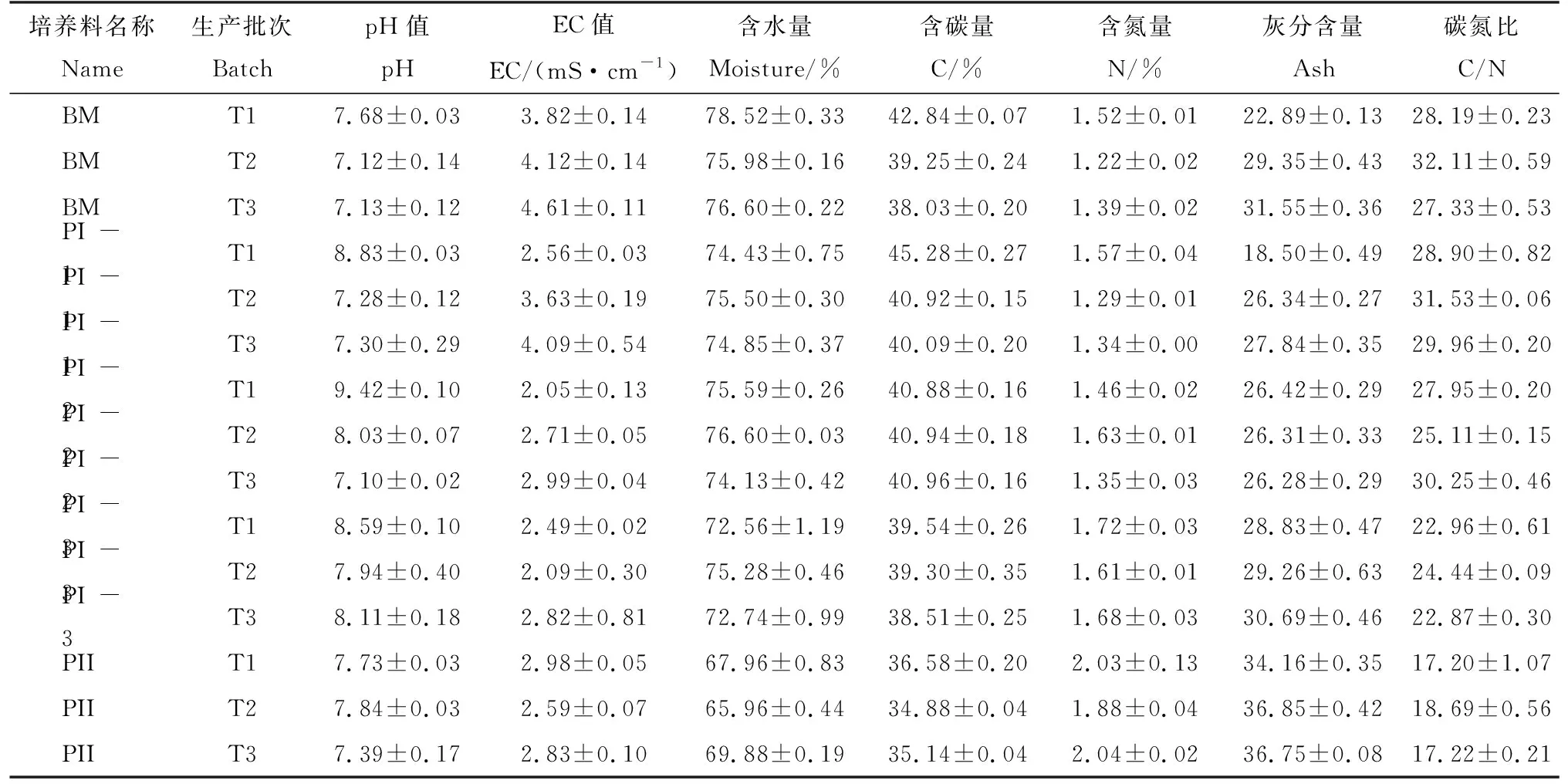
表1 3批次培养料堆肥时期理化性质Tab.1 Physical and chemical properties of three batches of samples during composting
表2是3批次培养料在栽培时期的理化性质。从菌丝长满(Filling)到第3潮菇(3rd)结束pH、含水量、含碳量、碳氮比呈逐渐下降趋势;EC值从菌丝长满(Filling)到第2潮菇(2nd)结束呈逐渐上升趋势。灰分含量从菌丝长满到第3潮菇结束逐渐上升,含氮量从菌丝长满的2.08到第3潮菇结束的2.03,整体变化不大。
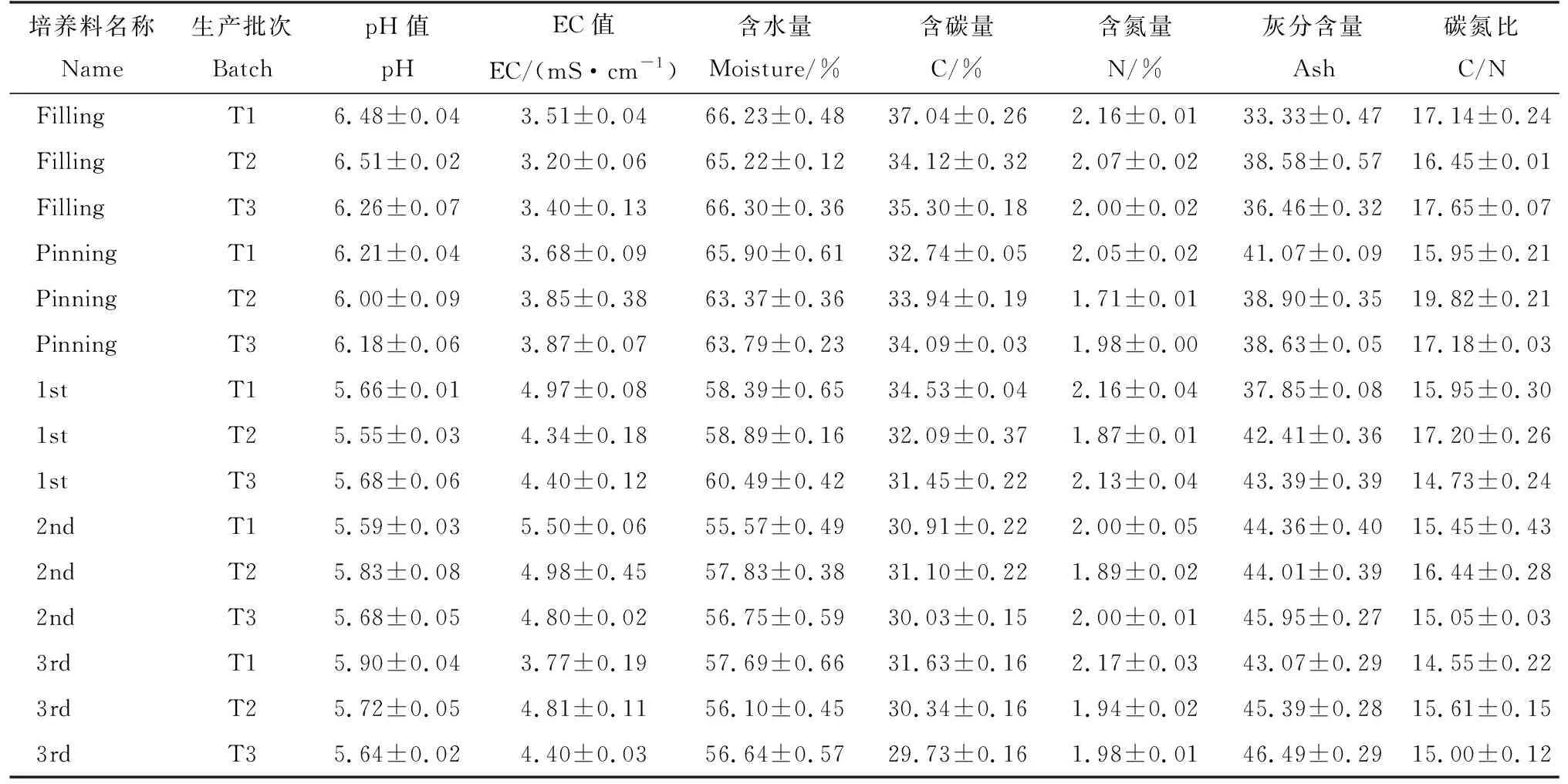
表2 3批次培养料栽培时期理化性质Tab.2 Physical and chemical properties of three batches of samples during cultivation
2.2 培养料木质纤维素相对含量变化
图1为培养料不同时期木质纤维素相对含量的变化。整体上看,3批次培养料不同时期的纤维素、半纤维素和木质素的相对含量变化趋势一致。在一次发酵时,半纤维素相对含量从19.64%降至16.67%,下降了2.97%;在二次发酵结束后相对含量降至10.93%,下降了5.74%,显著高于一次发酵;纤维素相对含量在一次发酵时维持在23.19%左右,二次发酵结束后降至15.73%,下降了7.46%,显著高于一次发酵;木质素经过一次发酵、二次发酵从16.12%增加至22.96%,相对含量显著增加,这可能与培养料的能量损失有关,表明堆肥时期木质素几乎没有被利用。从菌丝长满到第1潮菇时,半纤维素和纤维素相对含量有少量降低,半纤维素相对含量从7.71%降至4.73%,纤维素相对含量从13.96%降至10.31%;木质素相对含量从21.16%降至19.08%,三者均有所下降;出菇之后总碳水化合物的相对含量变化不明显,第3潮菇甚至有所增加,第2潮菇到第3潮菇半纤维素相对含量从3.87%增加到4.27%,纤维素相对含量从8.32%增加到8.37%,木质素相对含量从18.64%增加到18.98%。总体分析,纤维素和半纤维素在二次发酵时下降最多,木质素在堆肥时期几乎没有被利用,第1潮菇时3种成分均显著下降。
2.3 木质纤维素相关降解酶活性的变化
3批次培养料木质纤维素相关降解酶活性变化如图2所示。图2A、图2B均为降解纤维素的酶。羧甲基纤维素酶活性从建堆时到一次发酵结束都趋于相对稳定,一次发酵到二次发酵期间略有上升,上升至13.21 U/g,在二次发酵到原基形成略有
下降,下降至11.02 U/g,在第1潮菇时突然升高达到149.52 U/g,之后降低,第3潮菇降至69.42 U/g;α-纤维二糖酶活性在一次发酵时变化不大,在0.52 U/g左右,从一次发酵到二次发酵略有上升,从二次发酵结束到原基形成时又缓慢上升,从0.81 U/g升到1.56 U/g,第1潮菇时突然升高至13.30 U/g,之后缓慢降低,第3潮菇时降至7.75 U/g。由此可知,降解纤维素的2种酶在堆肥时期活性都相对平稳,栽培时期由于双孢蘑菇菌丝的旺盛生长,在第1潮菇时急速升高,以后逐渐下降,其中羧甲基纤维素酶的活性(11.02~149.52 U/g)要高于α-纤维二糖酶活性(0.37~13.30 U/g),是降解纤维素的主要酶类。
图2C、图2D、图2E分别为木聚糖酶、木糖苷酶、α-阿拉伯呋喃糖苷酶在蘑菇生长不同时期的活性变化,三者与培养料半纤维素的降解有关。木聚糖酶活性在堆肥时期维持66.09 U/g左右,在一次发酵的3次翻堆活性略有起伏,在二次发酵略有下降,从发菌时期开始活性显著提高,达140.43 U/g,不断升高,在第1潮菇达176.25 U/g,之后下降至145.73 U/g;木糖苷酶活性在一次发酵时变化明显,其中第2次倒仓时活性较高,由1.19 U/g升至1.92 U/g,之后呈下降趋势直至原基形成的0.55 U/g,又在第1潮菇时活性突然升高至5.07 U/g,之后一直下降,第3潮菇降至2.80 U/g;α-阿拉伯呋喃糖苷酶活性在堆肥时期的变化趋势与木糖苷酶活性相似,但是在一次发酵活性变化起伏较大,尤其是第2次倒仓时活性较高,达8.64 U/g,之后下降,二次发酵结束后降至5.27 U/g,在菌丝长满时突然升高至37.45 U/g,至原基时一直维持较高活性,之后在第1潮菇、第2潮菇、第3潮菇缓慢下降至34.52 U/g。由此可见,降解半纤维素的3种酶,在堆肥时期就具有一定的活性,其中木糖苷酶、α-阿拉伯呋喃糖苷酶在一次发酵第2次倒仓过程中活性较高。在出菇时期,3种酶在第1潮菇时活性都达到最高,其中α-阿拉伯呋喃糖苷酶活性在双孢蘑菇菌丝长满时就已经达到较高水平。3种酶中木聚糖酶的活性(58.65~176.25 U/g)要高于木糖苷酶活性(0.55~5.07 U/g)和α-阿拉伯呋喃糖苷酶的活性(2.29~39.60 U/g),这3种酶是分解半纤维素的主要酶类。
图2F为培养料不同时期漆酶活性的变化趋势,漆酶是降解木质素的酶。在堆肥时期漆酶几乎没有活性,在双孢蘑菇菌丝长满时活性突然升高至14.57 U/g,并在原基形成时保持较高活性,在第1潮菇时突然下降至0.70 U/g,之后又升高,第3潮菇升至14.03 U/g。3批次培养料各种酶活性的变化趋势基本一致。
从不同时期几种酶活性高低分析可知,在菌丝长满时分解半纤维素的木聚糖酶活性和α-阿拉伯呋喃糖苷酶活性就达到高值,之后持续在高水平,而分解纤维素的羧甲基纤维素酶和α-纤维二糖酶在第1潮菇时才达到较高的水平,漆酶在发菌时期和原基形成时较高,表明双孢蘑菇菌丝最先对半纤维素和木质素分解,而后对纤维素分解。
2.4 双孢蘑菇的产量和生物学效率
表3为3批次双孢蘑菇产量与生物学效率。双孢蘑菇总产量分别为20.08、20.28和25.98 kg/m2,生物学效率分别为52.05%、49.86%、71.03%。第3批次的生物学转化效率明显高于第1批次、第2批次。
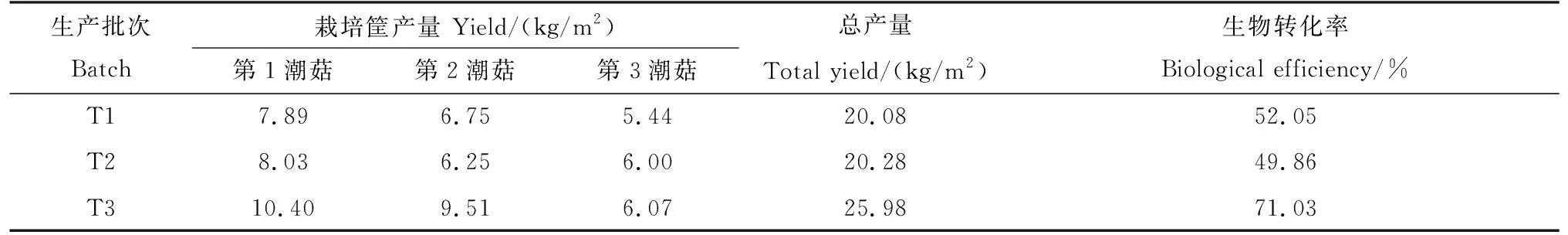
表3 蘑菇产量与生物学效率Tab.3 The yield and biological efficiency of mushroom
3 讨 论
堆肥质量是双孢蘑菇的生长和产量形成的基础。从3批次培养料的理化性质pH、EC、含氮量和含水量等都在双孢蘑菇培养料要求的范围内,与前人报道一致[17]。有研究表明,播种时(二次发酵结束)培养料的含氮量与蘑菇的产量呈现正相关,堆肥时期含水量是一个非常关键的因素[18]。该试验中第3批次生物学转化效率高于第1批次和第2批次,可能与其二次发酵的含水量、含氮量较高有关。
该试验3批次培养料不同时期的纤维素、半纤维素和木质素的相对含量变化趋势基本一致。堆肥时期在一次发酵时,半纤维素和纤维素相对含量降低3%左右,在二次发酵结束后下降更明显分别为7.46%和5.74%;出菇时期从菌丝长满到第1潮菇时半纤维素、纤维素、木质素相对含量均下降了3%左右,其中木质素相对含量在堆肥时期没有下降,在菌丝长满至原基形成时下降显著,与漆酶活性在这一时期达到高峰相吻合,表明双孢蘑菇菌丝生长时期分泌大量的漆酶,降解木质素。虽然双孢蘑菇属于典型的草腐型食用菌,但其分解木质素的能力在发菌时期较强,表明双孢蘑菇不仅利用纤维素、半纤维素,对木质素也很需要,这与Zhang[19]的研究结果一致。
不同时期木质纤维素的酶活性变化与木质纤维素各组分的降解有密切的关系。堆肥时期半纤维素、纤维素相对含量显著降低,此时期的羧甲基纤维素酶、α-纤维二糖酶、木聚糖酶、木糖苷酶、α-阿拉伯呋喃糖苷酶皆有一定的活性;堆肥时期木质素相对含量变化不明显,漆酶的活性比较低;出菇时期,在菌丝长满至第1潮菇时,双孢蘑菇菌丝体表现出较高的基质降解能力,尤其是漆酶活性自接种后激增,纤维素酶(羧甲基纤维素酶、α-纤维二糖酶)在原基形成时激增,半纤维素酶(木聚糖酶、木糖苷酶、α-阿拉伯呋喃糖苷酶)在菌丝长满时显著升高,第1潮菇时剧增,这一时期的木质素、纤维素和半纤维素相对含量显著下降。而第2潮菇之后随着各种酶活性的降低,纤维素、半纤维素、木质素的相对含量几乎没有下降,反而略有升高,Jurak等[16]的研究认为,由于酶活性的降低,木聚糖侧链残基积累影响纤维素、半纤维素的降解,该试验结果与其一致。该试验第1潮菇时产量最高,第2潮菇减产,第3潮菇产量最低,与Patyshakuliyeva等[20]的研究一致。试验3批次培养料中以第3批次二次发酵结束后的木聚糖酶活性(67.68 U/g)、羧甲基纤维素酶活性(15.16 U/g)、α-阿拉伯呋喃糖苷酶活性(8.35 U/g)、木糖苷酶活性(1.58 U/g)、α-纤维二糖酶活性(1.23 U/g)高于第1批次和第2批次,其产量表现最好,这与蔡盼盼[10]对10批次蘑菇二次发酵培养料酶活性与产量的相关分析结果一致。木质纤维素酶测定速度快、结果准确,以酶活性来评价二次发酵培养料的质量和预测产量具有一定的实用性,在试验测试条件较好的生产企业可以开发利用。
4 结 论
3批次培养料的理化性状在双孢蘑菇生长适宜范围,在堆肥时期纤维素、半纤维素被大量利用,以二次发酵时期其相对含量下降最显著,分别为7.46%和5.74%。在栽培时期,纤维素、半纤维素和木质素含量均下降了3%左右。各种降解酶在堆肥时期活性平稳,菌丝长满至第1潮菇时都急速升高,其中羧甲基纤维素酶的活性(11.012~149.52 U/g)远高于α-纤维二糖酶活性(0.37~13.30 U/g),是降解纤维素的主要酶类。木聚糖酶的活性(58.65~176.25 U/g)高于木糖苷酶活性(0.55~5.07 U/g)和α-阿拉伯呋喃糖苷酶的活性(2.29~39.60 U/g),是分解半纤维素的主要酶类。漆酶在堆肥时期几乎没有活性,菌丝长满至原基形成时活性高达14.57 U/g。3批次培养料中第3批次的各种酶活性、生物转化率明显高于第1批次和第2批次。各种酶活性与产量呈正相关。
