冬麦春播小麦对苗期干旱胁迫的生理响应
2020-12-28徐澜刘艳超安伟高志强
徐澜,刘艳超,安伟,高志强
(1.忻州师范学院生物系,山西 忻州 034000;2.山西省农科院玉米研究所,山西 忻州 034000;3.山西农业大学农学院,山西 太谷 030801)
小麦是我国重要的粮食作物、主要的商品粮和战略储备粮,其产量的稳定直接影响着粮食安全.冬春麦混播区是我国小麦生产的重要组成部分,该地域耕地资源丰富,光热资源充足,发展小麦生产极具潜力[1-2].然而冬春麦混播区春小麦生态类型复杂、品种较杂乱、优良品种资源缺乏,这些现实问题严重制约着当地小麦生产的发展.科学合理地引进、筛选、驯化优良小麦品种资源是保障小麦生产的重要措施之一.针对这一问题,高志强团队将长江中下游地区和西南冬麦区的优良冬播小麦品种引入到北方混播区(如忻定盆地)进行春播,即冬麦春播[3-5].冬麦春播是探索小麦品种资源更新、高效利用的一种新的途径.研究表明,经冬麦春播试验筛选得到的‘宁麦13’(2013产量:4 835.2 kg/hm2;2014产量:5 174.6 kg/hm2)等优良品种的产量显著高于当地春麦品种[3],增产潜力较大,对其在混播区的适应性、抗逆性进一步研究有重大意义.冬春麦混播区基本属干旱半干旱地区,近几年春旱频发,对于春小麦保苗、壮苗极为不利.小麦苗期是一个较为敏感的时期,是由异养转变为自养的关键时期,这时小麦的根系下扎还比较浅,容易发生旱害.因此,冬麦春播小麦的苗期耐、抗旱性表现是其引种成功与否的重要影响因素之一.
在长期的进化中,植物演化出一套有效抵御逆境的生理机制.通过渗透调节物质的增加,细胞质膜部分功能的丧失,自由基的产生和清除等因素综合作用来应对干旱胁迫[6-9].干旱胁迫抑制小麦生长发育,表现为植株生理特征的变化,其中可溶性蛋白含量、脯氨酸含量、质膜透性等是与抗旱性密切相关的生理指标.研究表明,干旱能显著影响小麦MDA的含量,且由于干旱胁迫造成大量的活性氧的积累,会引起细胞膜脂过氧化而产生MDA,导致MDA的含量升高并对细胞膜系统造成伤害,从而损害植物的生长[10-12].
抗逆性强弱受品种自身遗传特性的制约,筛选、驯化、培育抗旱性强的小麦品种是应对干旱气候的有效手段.为明确冬麦春播小麦品种在混播区的耐、抗旱性,本试验以前期(2013~2015年)混播区适应性引种筛选得到的高产小麦品种‘宁麦13’‘扬麦20’为材料,利用PEG 6000模拟干旱胁迫,研究了苗期不同干旱程度处理下2个品种的植株生长变化及生理生化响应,从而比较两品种的耐旱性强弱,为进一步研究冬麦春播小麦苗期抗旱生理机制提供参考,为混播区小麦安全生产提供备选品种及理论指导.
1 材料与方法
1.1 材料处理
以‘宁麦13’(南京)‘扬麦20’(扬州)为试验材料,将相同种子按距离1 cm间距种进同规格的育苗盆,每品种设置12盆,待小麦生长至二叶一心期,开始干旱胁迫处理.利用PEG 6000模拟干旱胁迫,用蒸馏水配置4个梯度的处理液,即PEG 6000浓度为0(T0)、8%(T1)、16%(T2)、24%(T3),各处理设置3次重复,分别在持续胁迫的第0天、第3天、第6天、第9天、第12天进行各项生理生化指标的测定,3次重复取平均值[12-13].
1.2 测定项目及方法
1.2.1 叶片面积 直尺测量展开叶的最长处和最宽处[14],取平均值,见式(1):
叶面积=最长×最宽×0.75
(1)
1.2.2 叶片相对含水量 在每个测量日同一时间,剪取‘宁麦13’‘扬麦20’相同叶位的叶片测定相对含水量(RWC)[15-16],见公式(2):
(2)
1.2.3 根和叶的相对电导率 剪取适量叶片,蒸馏水洗净,滤纸吸干水分,混匀,称取0.2 g,剪为0.5 cm左右相同规格的小段,加入20 mL蒸馏水,抽气、并使叶片完全浸入蒸馏水中,静置12 h;取适量小麦根系,用水清洗干净,蒸馏水冲洗数次,滤纸吸干水分,其余同叶片操作相同.静置12 h后,使用电导率仪测根和叶的初始电导率S1;之后将材料分别置于100 ℃水浴加热10 min,静置、冷却到室温,测电导率S2[17-18].见式(3):

(3)
1.2.4 可溶性蛋白含量 剪取‘宁麦13’‘扬麦20’的叶片适量,混匀,称取0.1 g,采用Tris-HCL缓冲溶液(pH=6.8)进行可溶性蛋白研磨提取,采用考马斯亮蓝法测定蛋白含量[3].以牛血清白蛋白(BSA)为标准品,在595 nm处绘制标准曲线:y=4.701 4x-0.004 6(R2=0.993 7).
1.2.5 丙二醛含量 剪取新鲜‘宁麦13’‘扬麦20’叶片适量,混匀,称取0.5 g,加入5 mL 10%三氯乙酸(TCA)溶液和少量石英砂进行研磨提取,提取液离心10 min,取上清液2 mL转移到试管中,加入2 mL 0.6%硫代巴比妥酸(TBA)溶液,摇匀,置于100 ℃水浴中加热15 min,取出冷却,上清液转移到离心管中,离心.取第二次离心后的上清液在450、532、600 nm处分别测定D值.对照用2 mL蒸馏水代替提取液.见式(4):
C=6.45×(A532-A600)-0.56×A450
(4)
上述式中C为丙二醛的浓度,μmol/L;A450、A532、A600分别为450、532、600 nm波长下的吸光值.
据式(5)计算‘宁麦13’‘扬麦20’叶片中丙二醛含量[19-21]:
MDA含量(μmol/g)=丙二醛浓度(μmol/L)×提取液体积(mL)/植物组织鲜质量(g)
(5)
1.2.5 抗旱系数 仿照文献计算抗寒系数的方法,使用公式(6)求出抗旱系数[22]:
抗旱系数(%)=胁迫后测定值/胁迫前测定值×100%
(6)
1.3 统计分析
应用Microsoft Excel 2010和SPSS 17.0软件进行数据处理和统计分析,采用单因素方差分析和多重比较(Duncan新复极差法)进行差异显著性检验.
2 结果与分析
2.1 苗期干旱胁迫对小麦叶面积的影响
由表1知,冬麦春播条件下苗期干旱胁迫第0天,不同处理(T0~T3)均表现为扬麦20的叶面积大于‘宁麦13’,即其叶面积大小存在品种遗传差异.T0处理下(CK)2种小麦长势良好,第12天时‘宁麦13’(8.69 cm2)‘扬麦20’(8.65 cm2)叶面积较接近,期间2个品种分别增长了64.3%、50.2%,对应T1处理下分别增长了58.2%、42.2%,干旱胁迫影响了叶片正常生长.T1处理下2个品种的叶面积在第9天接近,第12天‘宁麦13’超过‘扬麦20’;T2处理下2个品种的叶面积在第6天较接近,第9天‘宁麦13’超过扬麦20;T3处理下第9天时‘扬麦20’叶片已缺水萎蔫无法测定,‘宁麦13’叶片生长正常,但叶面积较同一时期T1、T2略小.同时,干旱胁迫(T1)第12天时2个品种叶面积均显著小于其CK(P<0.05),持续干旱胁迫(T1~T3)对‘宁麦13’、‘扬麦20’的叶片生长均造成不同程度的伤害;随着胁迫推进,‘宁麦13’逐渐缩小了与‘扬麦20’的叶面积差异,即‘宁麦13’的叶片在干旱逆境胁迫下生长力表现强于‘扬麦20’.T2、T3处理下对小麦叶片造成了不可逆的损害,抑制了小麦的正常生长.
2.2 苗期干旱胁迫对小麦叶片相对含水量(RWC)的影响
T0处理下,冬麦春播小麦品种‘宁麦13’‘扬麦20’的RWC在苗期基本稳定,随着干旱胁迫梯度(T1~T3)的增加,不同时期(9~12 d)2个品种的RWC均显著下降(P<0.05).T1处理下第6天‘扬麦20’的RWC首先降到80%以下,T2处理下第12天‘扬麦20’‘宁麦13’的RWC分别为30.05%、39.78%,T3处理下第9天已下降到30%左右,第12天‘扬麦20’叶片枯萎死亡,此时‘宁麦13’的RWC亦下降到19%.干旱胁迫12天时T1处理使‘宁麦13’‘扬麦20’的RWC分别下降11.3%、15.2%,T2处理RWC下降54.2%、63.6%.苗期持续干旱胁迫导致小麦叶片细胞水分亏缺甚至死亡,且干旱胁迫对‘扬麦20’叶片造成的不良影响更加严重.
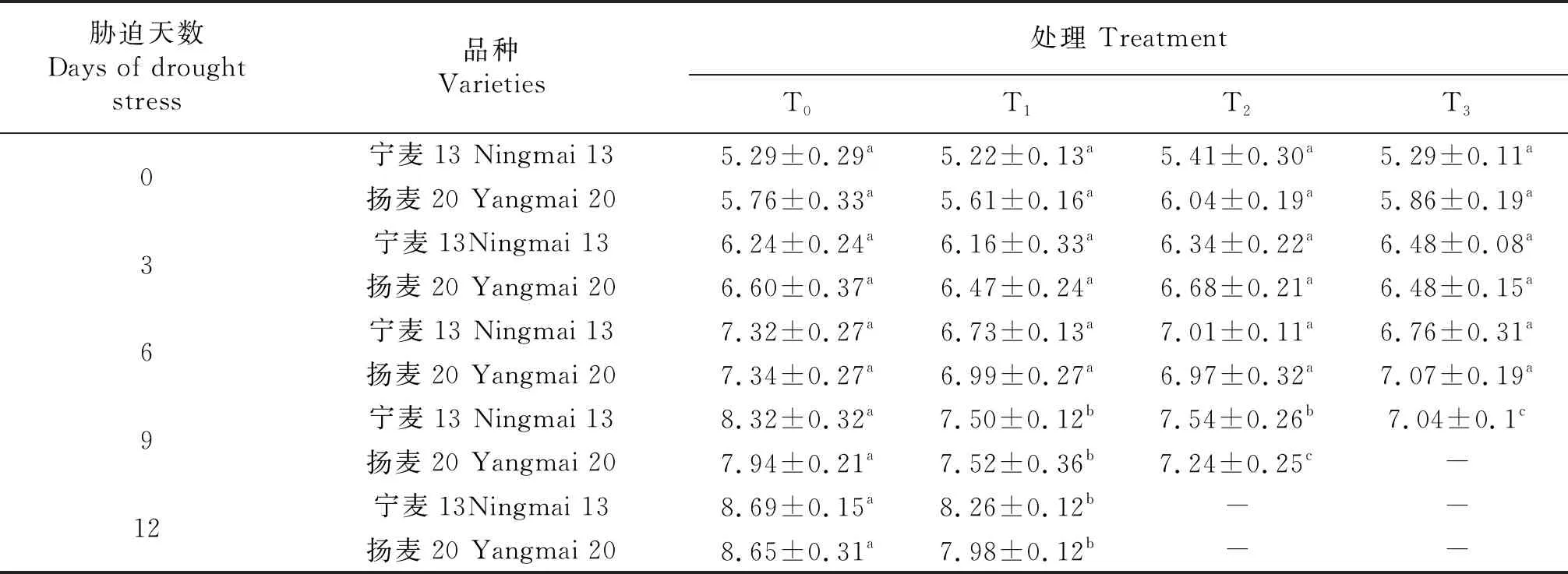
表1 苗期干旱胁迫对小麦叶面积的影响
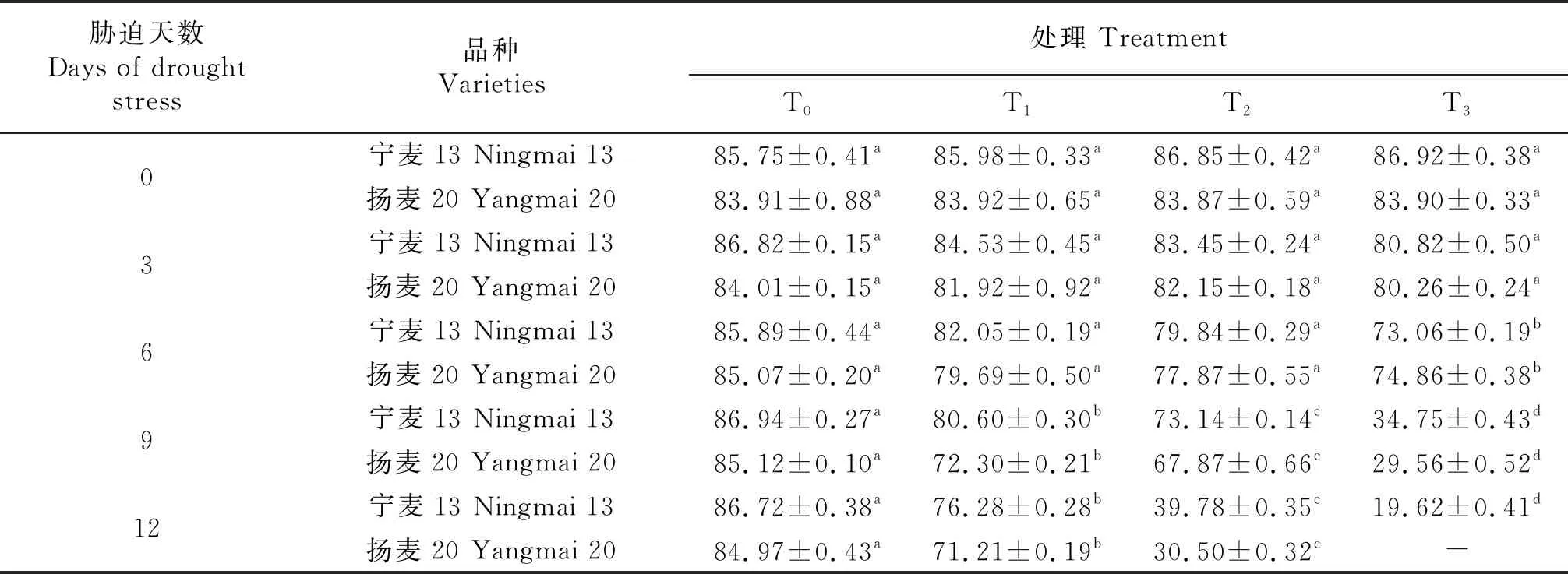
表2 苗期干旱胁迫对小麦叶片相对含水量的影响
2.3 苗期干旱胁迫对小麦根、叶电导率(REC)的影响
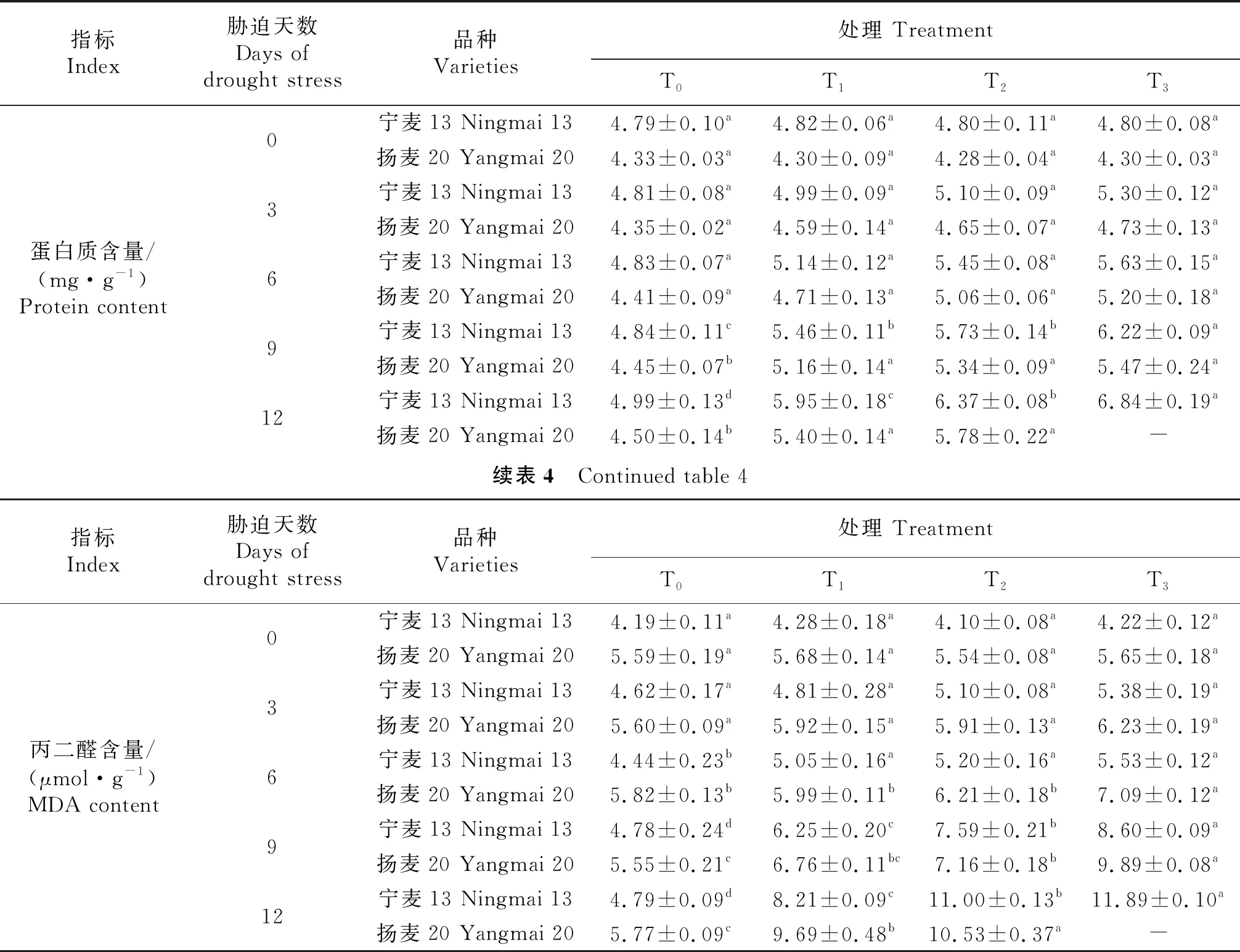
由表3知,冬麦春播条件下,‘宁麦13’‘扬麦20’的苗期相对电导率均为根系REC高于叶片REC两倍之多,T0处理下叶片REC为‘宁麦13’略高于‘扬麦20’,而根系REC是后者高于前者.‘扬麦20’在干旱胁迫第3天测定时叶片REC表现为T2、T3处理显著高于T0、T1,而‘宁麦13’各处理间差异不显著(P<0.05);‘扬麦20’在干旱胁迫第3天测定时根系REC表现为T1、T2、T3处理间差异不显著,但均显著高于T0;‘宁麦13’根系REC则为T2、T3处理显著高于T0、T1(P<0.05),即‘宁麦13’在T1处理下受干旱胁迫影响较‘扬麦20’为小.第6天及以后‘扬麦20’的根、叶REC在不同处理间差异均显著:T0 表3 苗期干旱胁迫对小麦根、叶相对电导率的影响Table 3 Effects of drought stress on REC of wheat root and leaf at Seedling Stage % 可溶性蛋白含量是衡量植物抗逆性的一个重要指标.由表4知,胁迫处理前‘扬麦20’的叶片可溶性蛋白含量比‘宁麦13’低14.2%,T0处理下随幼苗生长推进2个品种的叶片可溶性蛋白含量变化不明显.干旱胁迫第9天开始,2个品种不同程度地出现叶片可溶性蛋白含量的显著变化.胁迫第9天时,‘宁麦13’的叶片可溶性蛋白含量差异显著性为T0 表4 苗期干旱胁迫对小麦叶片可溶性蛋白含量、MDA含量的影响 MDA的产生会加剧细胞膜质的损伤,因此,在植物抗性生理实验研究中其含量是衡量植物抗逆性的一个常用指标.由表4知,正常情况下(T0)随幼苗生长推进两品种的叶片MDA含量变化不显著或略有增加.胁迫第6天,‘宁麦13’‘扬麦20’在T3处理下MDA含量分别为5.53、7.09 μmol/g;‘宁麦13’各干旱处理的MDA含量均显著高于CK,‘扬麦20’仅T3处理显著高于其它处理,T0、T1、T2之间差异并不显著;第9天时2个品种各处理间基本差异均显著,‘宁麦13’‘扬麦20’的MDA含量在干旱胁迫T3处理下比对照T0分别高82%、78%.可见,干旱胁迫加重小麦叶片细胞膜质过氧化,随胁迫程度及时间增加导致MDA含量显著增加. 根据各项抗旱指标的测定值,用公式求出单项指标的抗旱系数(表5),其中MDA含量的抗旱系数最大,可见冬麦春播小麦苗期对持续干旱胁迫比较敏感,在叶片含水量缓慢下降的同时细胞膜质过氧化程度已大大增加.除根REC外,其余各项指标均表现为‘宁麦13’的抗旱系数大于‘扬麦20’,其中蛋白质含量的抗旱系数2个品种基本一致.干旱胁迫对苗期‘宁麦13’、‘扬麦20’均产生显著影响,抑制了其叶片的正常生长的,叶片含水量显著下降,根、叶相对电导率、蛋白质含量、MDA含量均不同程度地增加,2种小麦在浓度≥24%的PEG 6000的胁迫处理下均不能正常生长;在浓度8%的PEG 6000处理下持续12 d,幼苗可以正常生长;但PEG6000浓度达16%时,幼苗正常生长只能坚持9 d.干旱胁迫下植株的生长是一个复杂的生理生化过程,2个品种对于试验中不同抗旱指标的响应亦有所差别. 表5 苗期干旱胁迫下小麦各指标的抗旱系数 植物抗旱性是一个涉及植物形态、生理生态特征及生理生化变化的综合反应,是由多种基因控制的复杂性状,利用单一指标评价具有很大的局限性和不确定性[22-23],评价的结果会具有较大差异.本试验根据文献及前期研究基础,选取相对电导率等5个关键指标来研究2种小麦对苗期干旱胁迫的响应,以消除单一指标的片面性,以期可以较为准确的评价其抗旱性.文献表明,小麦遭受干旱胁迫,其相对电导率增加,叶片相对含水量降低,植物体内的可溶性蛋白和丙二醛含量会增加[10,12,24];这与本试验研究结果所涉及的各项指标变化趋势基本一致. 相对电导率可以指示胁迫对植物的伤害率,‘宁麦13’的叶REC上升幅度较‘扬麦20’小,表明‘宁麦13’的叶片受到伤害较小,但‘宁麦13’根REC变化范围较‘扬麦20’根REC为大.由此推断,干旱胁迫从根传到叶的过程中,会诱导植物不同部位发生不同程度的生化反应,引起‘宁麦13’根系细胞离子渗漏严重,同时刺激其叶片产生保护作用,出现了根、叶不同部位对干旱胁迫响应的表现不尽一致,具体原因有待进一步研究.由于‘宁麦13’的叶片细胞质膜受到的伤害较小,因此在相同的干旱胁迫下其RWC高于‘扬麦20’;叶片含水量高,叶片的增长速度较快,试验亦证明‘宁麦13’的叶面积增幅较‘杨麦20’为大. 可溶性蛋白的含量亦是植物抗旱性的重要指标之一.植物组织里的可溶性蛋白是重要的渗透调节物质和营养物质,王保莉[20]等研究指出,干旱胁迫会导致植物新胁迫蛋白产生,新胁迫蛋白产生后能维持细胞内较低的渗透势,具有良好的渗透调节作用;与本研究结果一致:T1、T2处理下‘宁麦13’‘扬麦20’的叶片可溶性蛋白含量均表现为增加(分别为23.4%、25.5%和32.7%、35.0%),即冬麦春播小麦遭遇苗期干旱胁迫,均依靠合成更多的蛋白质来调节自身的渗透势,以此适应一定范围内的干旱胁迫.但候明[25]等研究指出,可溶性蛋白在短时间、低浓度的胁迫下会升高,随着时间和胁迫程度的增加,可溶性蛋白含量会降低;可以理解为,植物通过合成渗透调节物质蛋白质等来适应胁迫的能力是有限的,当胁迫强度超过植物自身可调节范围时,植物组织内可溶性总蛋白含量因遭受胁迫破坏会下降. 大量研究表明[25-28],干旱胁迫对植物体内细胞质膜造成不同程度的损伤,最终引起离子和渗透物质的失衡;同时,干旱胁迫下,大量的活性氧在植物体内积累,加剧质膜结构和完整性的破坏.膜脂过氧化产物MDA,会反过来降低抗氧化物的含量及抑制抗氧化酶的活性.因此,MDA含量可作为植物抗旱的重要生理指标之一.本研究结果亦表明,PEG 6000模拟的干旱胁迫下,小麦叶片MDA的含量均明显高于对照组,随着胁迫时间的延长,‘宁麦13’的涨幅较‘扬麦20’为大,MDA含量的升高表明干旱胁迫会造成酶保护反应和自由基生成的动态平衡失衡,进而造成膜脂过氧化的程度升高. 本试验选用‘宁麦13’(5 174.6 kg/hm2,4 835.2 kg/hm2)、‘扬麦20’(3 330 kg/hm2)为材料,该2个品种经多年冬麦春播引种试验证明,在忻定盆地栽培条件下适应性较强,具有稳产、多抗等优良性状,产量均高于当地春小麦品种[3].而且这2个品种亦是冬麦春播小麦的代表类型(发育进程、穗部性状等不同),苗期干旱胁迫T3处理持续9 d时,‘宁麦13’‘扬麦20’开始无法正常测定相关指标,叶片萎蔫甚至死亡,可见24%的PEG 6000干旱胁迫超过了小麦承受范围,没有实际意义.T1处理下的两种小麦基本能正常存活并且与T0处理下各指标差距较小,根据前文整体指标分析,冬麦春播小麦苗期的抗旱范围大约在0~16%的PEG 6000之间. 采用PEG 6000模拟干旱胁迫12 d之内,‘宁麦13’‘扬麦20’可能忍受干旱的存活范围的浓度为0~16%;苗期干旱胁迫不同程度地导致小麦叶片生长缓慢、含水量显著下降,根、叶相对电导率上升且根系表现更加敏感;同时,干旱诱导蛋白质含量增加:23.4%~35.0%;叶片MDA含量增加最为显著(抗旱系数为169.9%~229.6%),可作为监测小麦苗期干旱胁迫的首选指标.比较而言,‘宁麦13’的叶面积生长较快,膜脂过氧化程度低,细胞质膜受伤害程度也较小,苗期抗旱性更强,可作为忻定盆地冬麦春播引种的优选品种.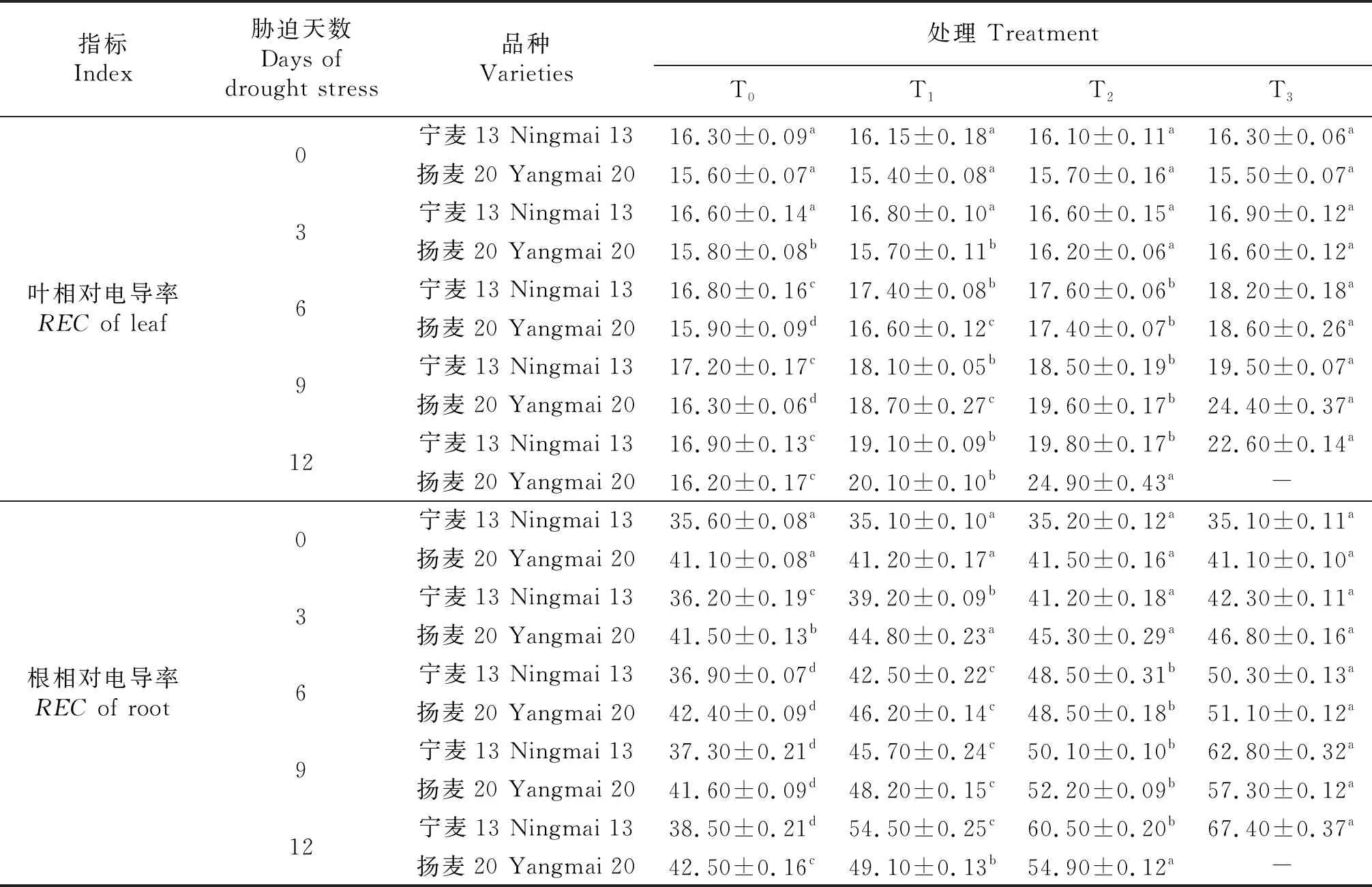
2.4 苗期干旱胁迫对小麦叶片可溶性蛋白、丙二醛含量的影响
2.5 苗期干旱胁迫下小麦各项指标的抗旱系数分析

3 讨论
4 结论
