‘黑珍珠’和‘飞弹’莲雾对催花处理的生理响应*
2020-12-15张绿萍金吉林
张绿萍,解 璞,金吉林,王 彬
(贵州省农业科学院果树科学研究所,贵阳 550006)
莲雾(Syzygium samarangense(Bi.)Merr.et Perry)系桃金娘科蒲桃属常绿果树。原产于马来半岛及安达曼群岛,果实形状优美、色泽鲜艳、口感清脆、清甜多汁。由于风味独特、市场需求量巨大、经济效益可观,莲雾已经成为东南亚地区的重要经济果树,也是我国重要的特色热带水果[1]。在市场需求驱动、国家政策引导和地方政府的大力支持下,近年来,栽培面积不断扩大,福建、海南、广东、广西、云南、四川、贵州等地均有栽培,经济效益很高[2]。
莲雾6—7月成熟的正季果由于采收季高温、多雨,导致果个小、着色不理想、风味和品质欠佳,又恰逢荔枝、龙眼、芒果等热带水果的盛产期,严重影响了其经济效益。通过产期调节,能使莲雾产期提早到冬春季,从而改善果实品质,提升市场竞争力和经济效益[2]。目前对莲雾产期调节的研究主要集中在栽培技术[2-6]和催花过程,尤其是遮阴处理期间的生理变化[7-9]。但以上研究均以易成功催花的‘黑珍珠’等粉红系品种为研究对象,以‘飞弹’等难催花的紫红系品种为研究对象的相关报道甚少,尤其未见关于催花处理过程中二者代谢差异的相关报道。本文以‘黑珍珠’和‘飞弹’为研究对象,比较二者催花处理后花芽分化情况、叶片碳水化合物和芽体内源激素含量的差异,探究莲雾花芽分化与碳水化合物、内源激素积累量及积累速度的关系,以及‘黑珍珠’产期调节成功而‘飞弹’失败的生理原因。以期为生产中不同莲雾品种产期调节提供理论依据和技术参考,并为进一步研究其花芽分化机理奠定基础。
1 材料与方法
1.1 试验材料
试验在罗甸试验基地莲雾种植场内进行,供试品种为易催花成功的‘黑珍珠’和不易催花的‘飞弹’,树龄6年,园地的土质属紫色壤土,肥力中等。
1.2 试验设计
选择已培育2批新枝、树冠茂密、新梢顶端叶片大多数由黄绿色转为绿色、长势相近的‘黑珍珠’和‘飞弹’莲雾各6株,2株为1组,重复3次。根据预试验结果,采用遮阴50 d+环割+喷催花剂方法进行催花处理,具体为7月5日采用遮光率为95%的遮阴网单株全面包裹的方式进行遮阴处理,8月5日环割,8月27日莲雾叶片呈八字形微微下垂时揭网,当日喷施催花药剂(48%乐斯本乳剂200倍液+80 mg/L NAA+10 g/L尿素+1.25 mL/L爱多收)。取样:遮阴期分别于7月5日、7月20日、8月4日及8月19日取样,药剂处理期自8月27日起,每3 d取样,到莲雾花芽出现(9月12日)为止。每株树在东、南、西、北4个方向的成熟枝梢上各取10片健康顶叶(共40片),洗净、擦干、杀青、烘干,用于可溶性糖、淀粉含量的测定。取顶芽及从上向下第2~5节芽(共180~210个),其中10个用于徒手切片观察,剩余芽用锡箔纸包好放入液氮中速冻,置于-70 ℃超低温冰箱保存,用于激素含量的测定。
1.3 开花情况调查
催花前在树冠中上部随机选定40个枝,东西南北每个方向10个,于9月25日调查其开花枝数,根据萌发花芽枝条数/调查枝条数计算开花枝条率;调查记录枝条上花芽数和总芽数,根据花芽数/总芽数计算枝条萌芽率;统计各研究对象的花穗数。
1.4 碳水化合物及激素含量的测定
可溶性糖含量采用蒽酮比色法测定,淀粉含量采用酸水解法测定,3次重复测定。激素含量采用高效液相色谱法测定,准确称取待测材料1.0 g,于弱光下液氮研磨,加入5 mL 80%冷甲醇,混匀后4 ℃避光浸提,过夜;浸提混合物3 500 r/min离心20 min,取上清液,沉淀中再加入2 mL 80%冷甲醇,混匀,4 ℃避光浸提1 h,离心,2次重复,合并上清液;上清液于40 ℃吹氮气至没有甲醇残余;剩余水相用等体积石油醚萃取脱色2次,弃去醚相;水相加入1~3滴甲酸调节pH值至3.0;再用等体积乙酸乙酯萃取3次,合并酯相,40 ℃吹氮气浓缩至干;最后加入0.5 mL甲醇溶解残质,用0.45 μm微孔滤膜过滤,进行高效液相色谱分析测试,3次重复测定。ZR、GA3、IAA、ABA含量测定的色谱条件为:高效液相色谱仪为美国Agilent公司产品,型号为1100 series,色谱柱:Eclipse XDB-C18(250 mm×4.6 mm,5 μm);流动相,甲醇∶0.075%乙酸水溶液=30∶70(v∶v);检测波长254 nm,流速1.0 mL/min,柱温35 ℃,进样量10 μL。所使用激素标样:ZR、GA3、IAA、ABA为美国Sigma公司产品。
2 结果与分析
2.1 催花处理后花芽萌发情况调查
从表1可知,‘黑珍珠’和‘飞弹’莲雾经催花处理后,均有花芽萌发,但6株‘黑珍珠’均出现花穗,未萌发大量叶芽,且正常开花结果;而‘飞弹’仅1株出现3串花穗,6株均在9月10日萌发大量叶芽,9月15日萌发大量新梢,最终3串花穗在开花前脱落。

表1 ‘黑珍珠’和‘飞弹’莲雾叶芽和花芽的萌发情况
2.2 喷药催花处理后花芽分化进程观察
由表2可知,8月27日揭网时,2个莲雾品种的芽体均未开始形态分化,喷施催花剂3 d后,可见为始分化期及分化末期芽体。9月2日,‘飞弹’可观察到叶芽生长,‘黑珍珠’可观察到花蕾原基分化。9月5日,‘黑珍珠’外观可见花芽萌出,花芽的形态分化进入花萼原基及花序生长期,‘飞弹’外观可见叶芽生长。9月9日,‘黑珍珠’可观察到花瓣原基及雄蕊原基。9月12日,‘黑珍珠’外观可见花序生长,花蕾分粒,花芽的形态分化进入雌蕊原基,部分可见完整小花,‘飞弹’可见叶芽生长(图版4)。

表2 催花处理后莲雾花芽分化进程
2.3 催花处理对‘黑珍珠’和‘飞弹’莲雾叶片碳水化合物的影响
2.3.1催花处理对‘黑珍珠’和‘飞弹’莲雾叶片可溶性糖含量的影响
遮阴后莲雾生长速度减缓,叶片逐渐转为深绿色,不再萌发新梢。由图1可知,‘黑珍珠’遮阴后,叶片可溶性糖开始积累,遮阴结束喷施催花药剂时达到峰值,随后随着花芽形态分化启动开始逐渐下降,进入雌蕊原基分化后略有回升。‘飞弹’遮阴初期可溶性糖含量略有下降,随后逐渐上升,遮阴结束喷施催花药剂时,叶片的可溶性糖含量与未遮阴时较为相近,无明显积累,喷施催花药剂后3 d有所上升,但与揭网时无显著差异,叶芽开始生长,随后6 d叶芽萌发,可溶性糖含量快速下降至最低值,此后出现小幅波动。另外,‘黑珍珠’叶片可溶性糖含量明显高于‘飞弹’,且每个时期含量差异明显,特别是遮阴期,随着遮阴时间的延长,二者叶片可溶性糖含量的差距越来越大。
2.3.2催花处理对‘黑珍珠’和‘飞弹’莲雾叶片淀粉含量的影响
由图2可知,‘黑珍珠’叶片淀粉的含量在遮阴前期略有下降,随后缓慢回升,揭网时略有积累,揭网喷施催花药剂后3 d快速上升,花芽形态分化启动,同期含量明显高于‘飞弹’,随后花蕾原基分化期略有下降,花萼原基、花瓣原基及雄蕊原基分化期持续上升,雌蕊原基分化期又有所下降,这可能是由于虽然花芽分化消耗大量可溶性糖,但揭网后光照变强,光合作用增强促进了淀粉的积累。‘飞弹’遮阴期,叶片淀粉含量持续缓慢降低,喷药后3 d仍小幅下降,随后随着叶芽生长不断上升。
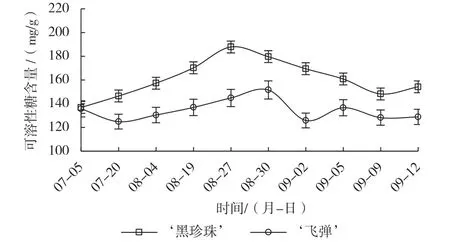
图1 催花处理对‘黑珍珠’和‘飞弹’莲雾叶片可溶性糖含量的影响
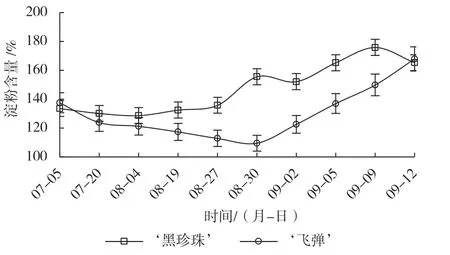
图2 催花处理对‘黑珍珠’和‘飞弹’莲雾叶片淀粉含量的影响
2.4 催花处理对‘黑珍珠’和‘飞弹’莲雾内源激素的影响
2.4.1催花处理对‘黑珍珠’和‘飞弹’莲雾ABA含量的影响
由图3可知,遮阴后‘黑珍珠’和‘飞弹’莲雾ABA含量均逐渐下降,揭网时二者的ABA含量相近。喷施催花药剂后,‘黑珍珠’的ABA含量快速上升,花芽始分化期和分化末期(8月30日)的含量约为未分化期(8月27日)的2.4倍,花蕾原基分化期(9月2日)达峰值,为未分化期的4倍,花萼原基、花瓣原基及雄蕊原基分化期快速下降,雌蕊原基分化期趋向平缓。‘飞弹’喷施催花药剂后呈“略有上升—快速上升—略有下降—快速下降”的变化趋势,8月30日的ABA含量仅比8月27日上升0.74 µg/g,不足‘黑珍珠’同期上升量的1.6%,随后3 d快速上升至峰值,与‘黑珍珠’同期达峰值,但其含量仅为‘黑珍珠’的1/2。
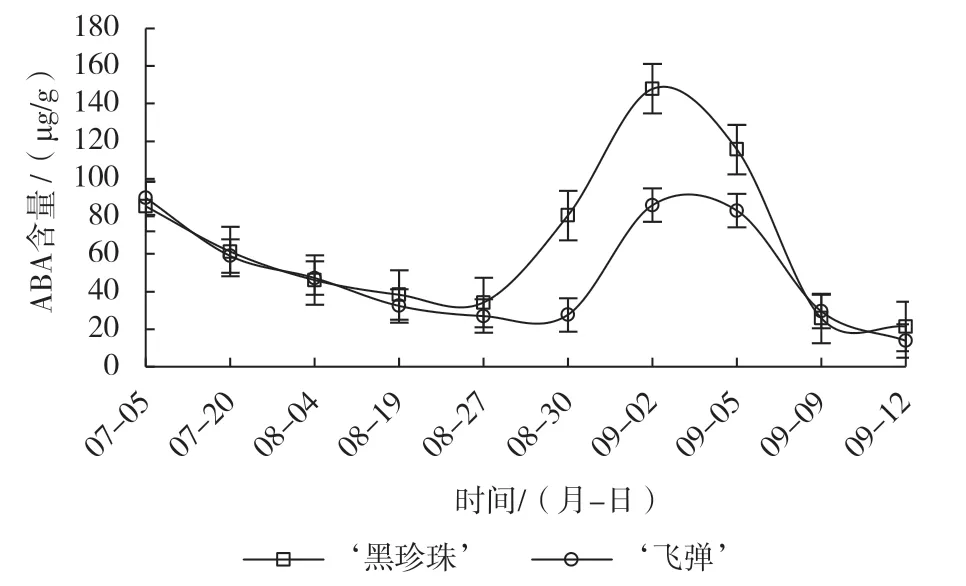
图3 催花处理对‘黑珍珠’和‘飞弹’莲雾ABA含量的影响
2.4.2催花处理对‘黑珍珠’和‘飞弹’莲雾IAA含量的影响
由图4可知,遮阴后芽体IAA含量持续下降,‘飞弹’的下降速率高于‘黑珍珠’,揭网时二者的IAA含量相近,‘飞弹’的IAA含量略低于‘黑珍珠’。喷施催花药剂后,‘黑珍珠’快速上升,花芽始分化期和分化末期(8月30日)达峰值,3 d的上升幅度为71.36 µg/g,其含量为未分化期(8月27日)的2.1倍,形态分化启动后,随着花芽分化,芽体的IAA含量逐渐下降,至雌蕊原基分化期趋向平缓。喷施催花药剂后,‘飞弹’呈“缓慢上升—急速上升—平缓上升—快速下降”的变化趋势。揭网喷施催花药剂后3 d,芽体IAA的上升幅度为20.68 µg/g,约为同期‘黑珍珠’上升幅度的1/4,叶芽萌动期(8月30日)的含量不足‘黑珍珠’花芽始分化和分化未期的1/2,随后叶芽开始萌发,芽体IAA上升至峰值,叶芽萌出(9月5日)后,芽体的IAA快速下降。
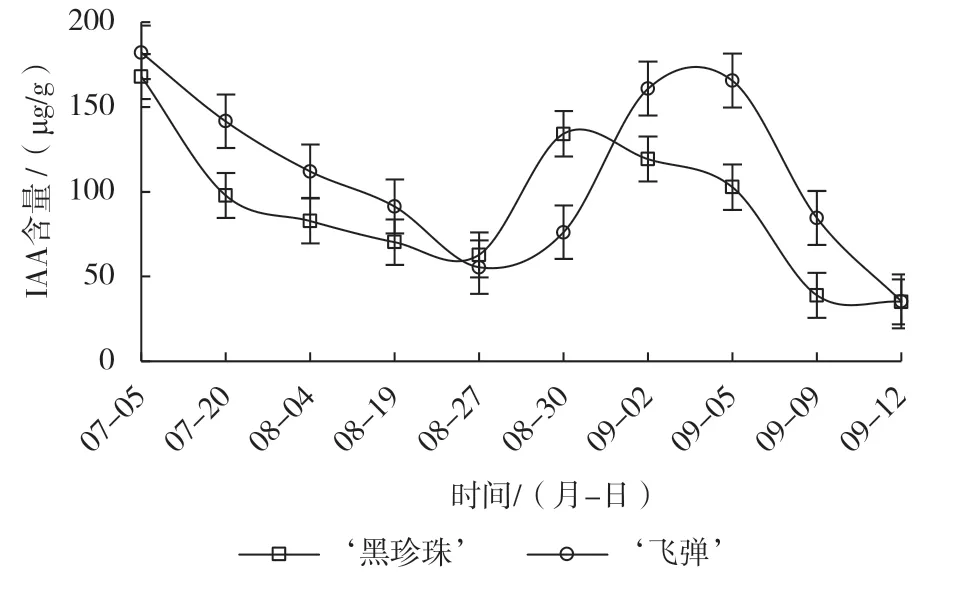
图4 催花处理对‘黑珍珠’和‘飞弹’莲雾IAA含量的影响
2.4.3催花处理对‘黑珍珠’和‘飞弹’莲雾GA3含量的影响
由图5可知,遮阴期‘黑珍珠’和‘飞弹’莲雾芽体GA3含量呈下降趋势,遮阴初期‘黑珍珠’的下降速率比‘飞弹’快,至7月20日起,‘黑珍珠’芽体的GA3含量明显低于‘飞弹’,揭网时,其GA3含量约为‘飞弹’的2/3。喷施催花药剂后,‘黑珍珠’芽体GA3含量的变化呈“上升—下降—趋于平缓”的趋势。花芽始分化和分化末期缓慢上升,3 d的上升幅度为35.36 µg/g,其含量约为未分化期的1.5倍,花蕾原基分化期(9月2日)上升至峰值,其含量约为未分化期的3倍,随后逐渐下降,至雌蕊原基分化期趋于平缓。‘飞弹’莲雾喷施催花药剂后3 d,叶芽萌动期芽体GA3含量快速上升至峰值,3 d的上升幅度为223.82 µg/g,是‘黑珍珠’同期增幅的6.3倍,其含量是叶芽未萌动期的3倍,是‘黑珍珠’同期含量的3倍。随后随着叶芽的萌发,芽体的GA3含量逐渐下降。
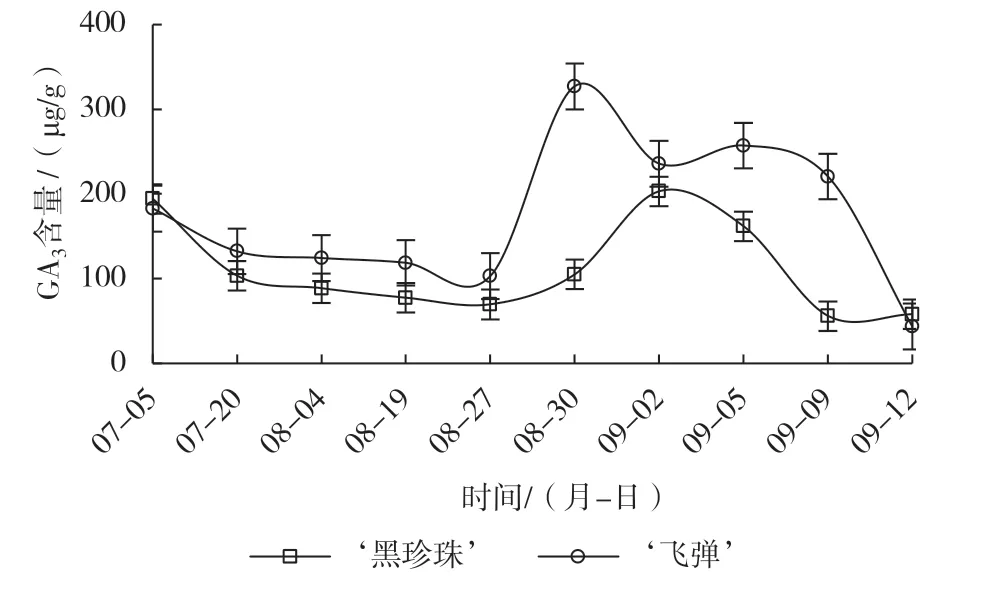
图5 催花处理对‘黑珍珠’和‘飞弹’莲雾GA3含量的影响
2.4.4催花处理对‘黑珍珠’和‘飞弹’莲雾ZT含量的影响
由图6可知,遮阴期‘黑珍珠’和‘飞弹’莲雾芽体ZT含量持续下降,下降速率相近,‘黑珍珠’ZT含量一直明显低于‘飞弹’,喷施催花药剂时,其含量约为‘飞弹’的1/2。喷施催花药剂后,二者ZT含量的变化趋势均为“上升—下降—略有上升—平缓下降”,但其上升幅度及上升维持的时间有明显差异。‘黑珍珠’喷施催花药剂后,花芽形态分化启动,芽体进入花芽始分化和分化末期,ZT含量快速上升,3 d的上升幅度为104.85 µg/g,其含量为153.06 µg/g,是未分化期的3.2倍,花蕾原基分化期(9月2日)达峰值,随后下降,趋于平缓。‘飞弹’喷施催花药剂后,叶芽萌动,芽体ZT含量上升至峰值,3 d的上升幅度为34.55 µg/g,不足同期‘黑珍珠’上升幅度的1/3,同期含量明显低于‘黑珍珠’,随着叶芽的萌发,芽体ZT含量逐渐下降后趋于平缓,并且同期含量均明显低于‘黑珍珠’。
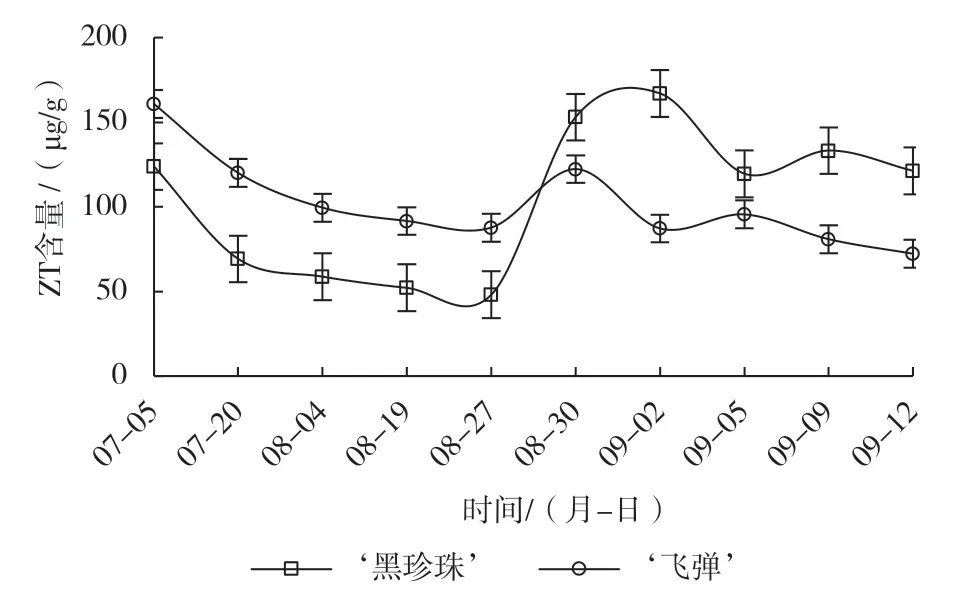
图6 催花处理对‘黑珍珠’和‘飞弹’莲雾ZT含量的影响
3 讨论与结论
莲雾产期调节的关键是抑制营养生长使其转入生殖生长,遮光处理,结合环剥、断根、浸水等栽培措施,控制营养生长,调整树势,再在花芽分化期配合化学药剂促进抽生花芽[4]。郑加协等研究指出,‘粉红种’莲雾仅进行遮光、环割、断根等栽培措施处理,植株没有花穗或者仅有5~10串花穗。栽培措施处理结束后喷施催花药剂,植株的花穗数可达200串,表明喷施催花药剂可显著提高树体的花芽分化率[2]。本试验结果表明,栽培措施处理结束时‘黑珍珠’和‘飞弹’仍处于未分化期,喷施催花药剂后3 d,‘黑珍珠’花芽发端,‘飞弹’叶芽萌动,可见喷施催花药剂后3 d可能是植株对药剂催花关键响应时期,也可能是莲雾花芽诱导和发端的临界期。这与章希娟等指出的揭网喷施催花药剂后的10 d为莲雾从营养生长转向生殖生长的关键时期的结果相对一致[8]。‘飞弹’莲雾催花失败与喷施催花药剂后3 d树体没有达到其花芽发端生理条件密切相关。
植物从营养生长转向生殖生长的过程中,植株体内新陈代谢旺盛,并产生着一系列的生理生化变化。碳水化合物既是结构物质又是能量提供者,其积累与花芽分化密切相关[10]。庞夫花等认为草莓花芽作为“库”,其分化和形态建成与碳水化合物密切相关[11]。罗羽洧认为无花果花芽分化与碳水化合物含量有一定的关系[12]。本试验结果表明,莲雾叶片的可溶性糖含量与其花芽的孕育和发端密切相关,足够的可溶性糖可保证其花芽分化的启动。这与张丽梅等[7]和郑加协等[2]的研究结果一致,莲雾喷施催花药剂前须积累足够的碳水化合物才能有效催花。
淀粉作为植物体内主要的贮藏性营养物质,花芽是否分化与其含量也密切相关,尤其是花芽发端和启动期,高水平的淀粉含量能有效保证其花芽分化的启动及进程。这与全振炫等[9]的研究结果一致,‘黑珍珠’莲雾遮阴处理控制营养生长,促进顶芽淀粉粒的积累,储备一定的碳水化合物确保顶芽花芽成花。‘飞弹’莲雾催花失败,与遮阴期不能积累足够的可溶性糖,持续消耗淀粉,喷施催花药剂后3 d仍无法有效积累相关,也就是遮阴、环割、断根等栽培措施不能有效促进其碳水化合物积累密切相关。其原因可能是‘飞弹’比‘黑珍珠’不耐弱光,在90%~95%的遮光率下光合产物不能有效合成,正如不耐弱光的葡萄品种‘夏黑’在设施促早栽培出现冬芽形成花芽不足,而且花芽质量差的现象[13-14]。如果选择遮光率较低的遮阴网或其他抑制营养生长的栽培措施,抑制植株营养生长的同时,保证其较强的光合作用,促进碳水化合物的积累,从而保证‘飞弹’莲雾喷施催花药剂后花芽分化启动时碳水化合物这一重要的物质基础,有待于进一步探索。为保证品质优良的‘飞弹’莲雾催花成功,可参考‘印尼红’莲雾的产期调节技术,采用遮光率为75%遮阴网单株包裹遮阴[15]。
植物花芽分化是一个复杂的过程,受一系列环境和内源因子调控,有研究认为,果树从叶芽转向花芽发育的不可逆过程需要植物激素在含量上的变化波动进行调控[16]。本试验中发现,遮阴期‘黑珍珠’和‘飞弹’莲雾芽体内源激素逐渐减少,揭网喷施催花药剂时ABA、IAA、GA3、ZT的含量相近,揭网喷施催花药剂后3 d,二者的各种内源激素变化和含量差异明显,结合花芽分化的进程和‘黑珍珠’‘飞弹’催花结果,表明遮阴期内源激素的含量和变化趋势与莲雾花芽是否分化的关系不密切,揭网喷施催花药剂后3 d内源激素的含量和变化趋势与莲雾花芽是否分化的关系极为密切。激素是花芽分化的关键内源因子之一,各个激素都会对花芽分化起一定的作用并呈现一定的规律[17-18]。多数学者认为,高水平的ABA、ZR,低水平的GA3有利于花芽分化的启动,而IAA在果树花芽分化中的作用尚无定论,既有高含量促进花芽分化的报道,也有低含量促进花芽分化的证据。‘首红’苹果花芽生理分化过程中,高水平的ABA、ZR和低水平的IAA、GAs互相作用促进了成花[19];刺梨花芽生理分化需要高浓度的ZRs与低浓度的IAA、GAs协同作用[20];杨梅花芽生理分化期叶片ABA、IAA含量上升,GA3含量下降是其花芽分化启动所必需的[21]。‘黄金杏’花芽生理分化期芽体ABA含量不断上升、IAA含量不断下降,ZR维持在相对稳定的较低水平[10]。‘宁玉’草莓花芽分化需要高水平的ZR、ABA和低水平的 GA3、IAA[11]。本试验结果表明,莲雾花芽分化与芽体内源激素积累速度和积累量均密切相关,高水平的ABA、IAA、ZT,低水平GA3,以及ABA、IAA、ZT的快速积累有利于其花芽分化的发端和形态分化的启动。这与高水平的ABA、ZT,低水平的GA3有利于‘粉红种’莲雾反季节催花后成花的结论一致[8]。为保证‘飞弹’莲雾催花成功,可考虑改变催花药剂的配方,结合遮阴期喷施多效唑和促花灵,环剥时环剥口涂抹促花灵[15],从而促进喷施催花药剂后芽体ABA、IAA及ZT的快速积累,抑制GA3的积累,处理的浓度、方法有待于进一步探索。
莲雾催花成功与否首先是通过遮光、环割、断根等栽培措施是否有效促进碳水化合物积累,以保证莲雾花芽孕育、启动、分化的重要物质基础。其次是喷施催花药剂后3~10 d花芽发端的关键期。外源药剂可促进芽体激素ABA、IAA、ZT含量快速积累,并抑制GA3的含量增长,以此促进花芽形态建成。
