草鱼IRE1-like基因的克隆、组织表达及其对微囊藻毒素-LR的响应
2020-12-10吴富盛阮记明隗黎丽
刘 林 何 丽 吴富盛 阮记明 周 颖 隗黎丽
(江西农业大学动物科学技术学院, 南昌 330045)
内质网承担着蛋白质的折叠、组装和转运。在蛋白质折叠过程中, 由于各种原因(DNA损伤、化学诱导和病毒感染等)使得未折叠蛋白或错误折叠蛋白增多[1], 未折叠蛋白反应(Unfolded protein response, UPR)可通过调整内质网折叠蛋白的数量和加强内质网的折叠能力, 减少未折叠蛋白或错误折叠蛋白的进一步产生, 从而减缓内质网应激使细胞存活[2,3]。其中, 需肌醇酶1(Inositol-requiring enzyme 1,IRE1)是参与UPR的重要分子。当未折叠蛋白在内质网腔中积累时,IRE1可通过信号转导调节特定基因和蛋白的表达[4]。当内质网压力过于严重不能被有效清除时,IRE1可引起细胞质内钙离子紊乱并诱导凋亡相关信号分子的表达[5,6], 同时,IRE1自身也能降解内质网中的错误蛋白, 从而释放内质网的压力[7]。在20世纪90年代, 哺乳动物的IRE1基因cDNA序列已相继被克隆[8—10], 而水产动物中IRE1基因的研究较晚, 近几年才在黄颡鱼(Pel-teobagrus fulvidraco)[11]和南美白对虾(Litopenaeus vannamei)[12]中克隆并进行研究。
草鱼(Ctenopharyngodon idella)作为我国四大家鱼之一, 养殖面积非常大。近年来, 随着养殖环境的恶化, 草鱼养殖水体易暴发蓝藻水华产生次生代谢产物微囊藻毒素(Microcystins, MCs), 微囊藻毒素-LR(Microcysitin-LR, MC-LR)作为MCs家族中的一员, 具有非常强的肝毒性[13]。有报道表明MC-LR可激活细胞内质网应激通路, 活化核转录因子-κB(NF-κB)、肿瘤坏死因子(TNF-α)和干扰素-α (IFN-α)的表达而诱导炎症反应、细胞凋亡和肿瘤[14]。在对MC-LR处理后的草鱼肝脏转录组进行测序分析的基础上[15], 筛选得到了草鱼IRE1-like基因的EST序列。本研究根据EST序列进一步克隆出草鱼IRE1-like基因cDNA全长序列, 分析了基因结构、在不同组织中的表达模式及不同剂量MC-LR诱导草鱼不同时间后的表达变化, 为进一步深入了解IRE1基因在鱼类响应MC-LR中的作用奠定基础。
1 材料与方法
1.1 实验鱼的处理
本实验所用的草鱼(22.13±2.17) g购于江西省南昌神龙渔业公司, 在实验室暂养两周后, 随机取3尾鱼用于草鱼IRE1-like基因组成型表达的分析,测定的组织包括头肾、肾、皮肤、肠、心脏、鳃、脾脏、肌肉及肝脏。随后, 将剩余的草鱼随机分组进行MC-LR染毒试验, 包括对照组和实验组,每个组设置三个重复组。试验前, MC-LR(纯度≥95%)粉末用甲醇溶解成1 μg/μL的母液保存于–20℃, 使用前用0.8%生理盐水稀释成25 μg MCLR/kg体重(25 μg MC-LR/kg body weight, 25 μg MC-LR/kg BW)和100 μg MC-LR/kg BW两个剂量。实验组草鱼经腹腔注射染毒, 注射量为0.1 mL/尾; 对照组每尾草鱼经腹腔注射等量的0.8%的生理盐水。在注射MC-LR 24h和96h后, 分别从实验组和对照组中各取6尾鱼分离肝脏, 置于液氮中保存。为避免应激反应, 在注射以及取样时, 所有鱼均用100 μg/mL的间氨基苯甲酸乙酯甲烷磺酸盐(MS-222)进行麻醉。
1.2 实验试剂
MS-222为Sigma公司产品, RNA提取试剂盒为Invitrogen公司产品, 检测表达分析的逆转录试剂盒RevertAidTMFirst Strand cDNA Synthesis Kit以及荧光定量PCR的SYBR Green Real-time PCR Master Mix均为Promega公司产品, 逆转录试剂盒、凝胶纯化回收试剂盒、Taq酶及Maker等购自TaKaRa公司。其他试剂如氯仿、无水乙醇和异丙醇等为中国国药分析纯产品。
1.3 核酸的提取及cDNA的合成
取出保存在液氮中的样品, 采用Trizol试剂盒提取总RNA, 具体提取方法参考试剂盒的说明书进行。用于检测不同组织表达以及MC-LR诱导表达的第一链cDNA用RevertAidTMFirst Strand cDNA Synthesis Kit试剂盒进行反转录合成, 用于RACE扩增的cDNA按照Super SMARTTMPCR cDNA Synthesis Kit操作手册的方法进行反转录, 分别合成3′RACE-Ready cDNA和5′ RACE-Ready cDNA。
1.4 草鱼IRE1-like基因cDNA全长的克隆
根据转录组测序数据获得的草鱼IRE1-like序列,利用Primer Premier 5.0设计引物如表 1, 由上海生工生物工程股份有限公司合成, 以草鱼肝脏cDNA为模板进行PCR扩增, 并对所获得片段进行胶回收和测序,比对分析确定获得的序列为草鱼IRE1-like基因中间序列。再根据这一序列设计上游和下游的特异性嵌套引物(表 1), 按照SMARTTMcDNA Amplification Kit (Clontech)说明书推荐的反应体系及反应条件进行3′ RACE和5′RACE扩增。PCR反应程序及条件等参照实验室发表文献设置[16], 分别获得3′-和5′-末端序列, 再与中间序列拼接得到全长cDNA序列。

表 1 本文所用引物序列Tab. 1 Primers used in the study
1.5 草鱼IRE1-like基因序列分析及系统发育树的构建
利用生物信息学在线工具对草鱼IRE1-like基因进行分析, 所用分析在线软件及其网址见表 2。系统发育树则采用Mega7.0软件中的NJ法进行构建, 在建树前氨基酸序列的比对用ClustalW1.81软件分析。
1.6 草鱼IRE1-like基因组织分布特征分析
根据获得的草鱼IRE1-like基因设计荧光定量引物IRE1-like-QF/R (表 1), 用于检测草鱼IRE1-like基因的组织分布。按照前述选择的3尾草鱼, 取头肾、肾、皮肤、肠、心脏、鳃、脾脏、肌肉及肝脏提取RNA反转录的cDNA模板, 使用Real-time quantitative PCR (qRT-PCR)方法检测草鱼IRE1-like基因的表达水平。qRT-PCR采用CFX96 Touch™Real-Time PCR Detection System, 20 μL反应体系包括: 10 μL SYBR Green Real-time PCR Master Mix,5.0 μL cDNA模板(50倍稀释的cDNA), 上下游引物各0.5 μL (20 μmol/L)和4.0 μL ddH2O。反应程序为: 95℃变性5min; 95℃ 10s, 58℃ 15s, 72℃ 20s,40个循环后, 72℃延伸5min。以草鱼β-actin基因作为内参基因进行校正, 采用2–ΔΔCt法计算IRE1-like基因的相对表达量。

表 2 生物信息学分析所用软件及网址Tab. 2 Software and websites used for bioinformatics analysis
1.7 微囊藻毒素-LR诱导草鱼IRE1-like基因的表达分析
分别提取25和100 μg/kg BW 剂量MC-LR诱导草鱼24h和96h后的RNA样品, 按前述方法反转录合成cDNA进行qRT-PCR, 检测MC-LR诱导草鱼肝脏IRE1-like基因在不同时间点的表达情况。反应体系、反应程序及数据分析如1.6所述。
1.8 数据分析
实验数据均以平均值±标准差表示, 其中, 草鱼IRE1-like基因在不同组织中的相对表达量采用One-way ANOVA (SPSS 16.0)分析, MC-LR对草鱼IRE1-like基因表达的影响采用多因素方差分析进行统计学检验(SPSS 16.0), 统计学显著性水平设定P<0.05表示差异显著。
2 结果
2.1 草鱼IRE1-like基因全长cDNA序列特征分析
根据草鱼转录组测序得到一段长约3000 bpIRE1-like基因的序列, 设计引物对该段序列进行验证并在NCBI上进行BLAST分析, 进一步确定为草鱼IRE1-like基因cDNA的中间序列, 随后通过RACE法扩增草鱼IRE1-like基因的5′和3′末端序列,分别获得了长度为309和549 bp大小的产物。经序列拼接获得草鱼IRE1-like基因cDNA全长序列, 其在NCBI的GenBank登录号为MG797683。草鱼IRE1-like基因全长3595 bp, 包括5′非编码区和3′非编码区, 分别为111和391 bp, 开放阅读框(Open Reading Frame, ORF)为3093 bp。对其编码氨基酸序列预测分析, 发现草鱼IRE1-like编码1030个氨基酸, 分子量为116.24 kD, 理论等电点为6.26。采用SignalP4.1在线分析表明草鱼IRE1-like蛋白存在信号肽, 最可能的切割位点在17和18氨基酸残基(ITA-II)之间。采用Phyer2跨膜结构预测发现草鱼IRE1-like蛋白在第755和第770之间的氨基酸存在跨膜螺旋结构。在InterPro上分析序列发现, 草鱼IRE1-like蛋白包含三个超家族, 分别为WD40/YVTN repeat-like-containing domain superfamily (9—314 aa)、Quinoprotein alcohol dehydrogenase-like superfamily (26—236 aa)、Protein kinase-like domain superfamily (572—854 aa)和KEN domain superfamily(836—976 aa), 此外, 还发现草鱼IRE1-like蛋白有5个Pyrrolo-quinoline quinone beta-propeller重复结构(34—66、117—149、155—187、199—230和286—317 aa)。在NCBI数据库中, 草鱼IRE1-like蛋白序列同源搜索的结果显示, 39—307、569—834、837—915的氨基酸残基位置上分别存在管腔结构域(Luminal)、丝/苏氨酸蛋白激酶催化结构域(STKc)及核糖核酸内切酶结构域(RNase)三个保守结构域。通过NetOGlyc4.0分析(分值大于0.5的预测为糖基化位点), 发现草鱼IRE1-like蛋白存在共56个糖基化位点, 35个为丝氨酸残基, 21是苏氨酸残基; 利用NetPhos 3.1对草鱼IRE1-like蛋白进行磷酸化位点预测分析, 发现120个磷酸化位点, 其中丝氨酸残基72个, 苏氨酸残基42个, 酪氨酸残基6个。
2.2 草鱼IRE1-like氨基酸同源性与系统进化分析
草鱼IRE1-like氨基酸与斑马鱼(Danio rerio)IRE1-α、金线鲃(Sinocyclocheilus grahami) IRE1-like和鲫(Carassius auratus) IRE1-like的相似性和一致性非常高, 分别达到了95%—90%和97%—92%,与其他物种的相似性和一致性分别为86%—74%和91%—83%, 而与人(Homo sapiens)和小鼠(Mus musculus)IRE1-β的相似性则较低, 仅为48%—49%。应用MEGA7.0软件Neighbor-joining(NJ)法构建的系统进化树表明人和小鼠的IRE1-β单独聚为一支,人和小鼠以及爬行类和鸟类的IRE1聚为一支, 鱼类的IRE1及IRE1-like聚为一支, 其中草鱼IRE1-like与斑马鱼的IRE1-α亲缘关系最近(图 1)。
2.3 草鱼IRE1-like基因组织分布特征分析
对草鱼头肾、肾、皮肤、肠、心脏、鳃、脾脏、肌肉及肝脏9种组织中草鱼IRE1-like基因的表达进行了qRT-PCR研究, 发现草鱼IRE1-like基因在所有检测的组织中均有表达, 其中在肝脏中的表达量最为丰富, 其次为肌肉、脾脏、鳃和心脏等, 在头肾中表达量相对较低(图 2)。以相对表达量最低的头肾作为参照进行One-way ANOVA分析, 结果发现肝脏、肌肉、脾脏、鳃、心脏及肠道中草鱼IRE1-like的表达量显著高于头肾中的表达量(P<0.05)。
2.4 微囊藻毒素-LR对草鱼肝脏IRE1-like基因水平的影响
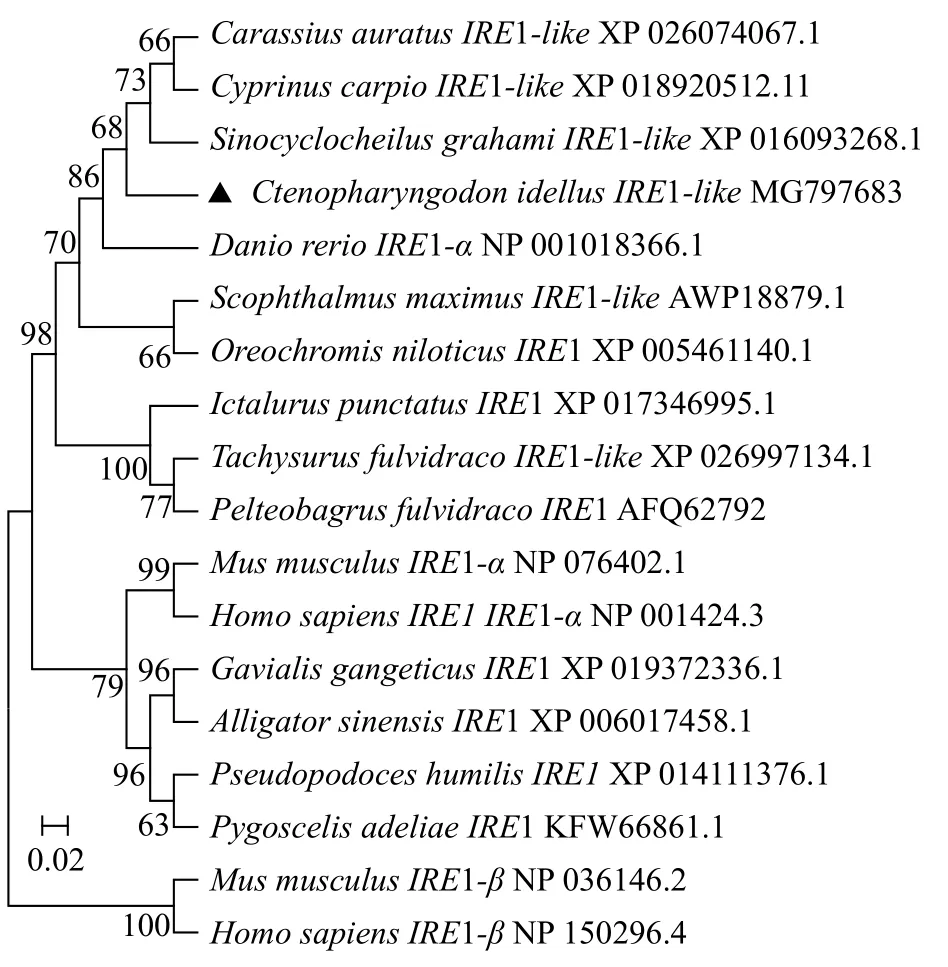
图 1 草鱼及其他脊椎动物IRE1基因的系统进化树Fig. 1 Phylogenetic tree of IRE1 of grass carp and other species
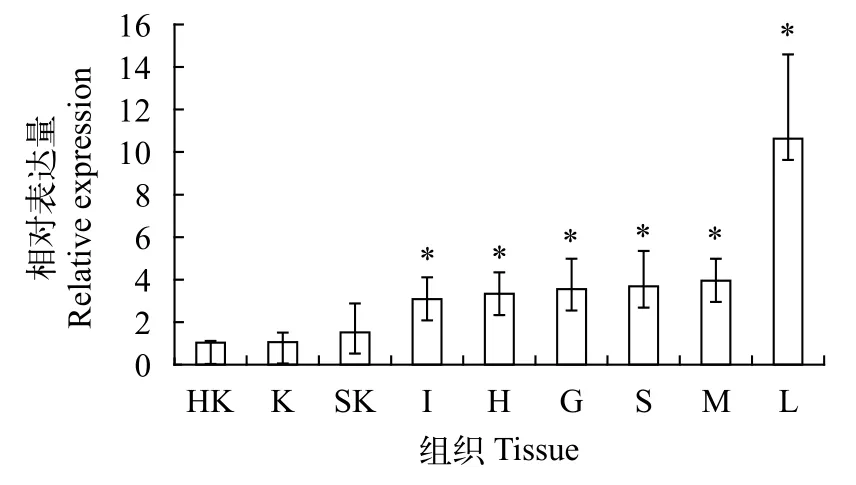
图 2 草鱼IRE1-like基因的组织特异性表达分析Fig. 2 Tissue-specific expression of IRE1-like in grass carp
如图 3所示, 在25 μg/kg BW MC-LR诱导后, 草鱼肝脏IRE1-like基因相对表达量在24h后显著升高(P<0.05), 而在96h表达上调但与对照组差异不显著(P>0.05); 100 μg/kg BW MC-LR诱导草鱼24h后,IRE1-like基因相对表达量升高(P>0.05), 而随着时间的延长, 草鱼肝脏中IRE1-like的表达量较对照组显著上升(P<0.05)。
3 讨论
3.1 IRE1-like基因cDNA全长的克隆及生物信息学分析
本研究克隆了草鱼IRE1-like基因的cDNA全长序列, 编码区为3093 bp, 共编码1030个氨基酸, 含有Luminal、STKc和RNase三个保守结构域, 这与黄颡鱼[11]IRE1和团头鲂[12]IRE1的结构域相同。在之前对人类和哺乳动物的研究发现,IRE1是双官能团单跨膜蛋白, 包括Luminal、STKc及核糖核酸内切酶区RNase, 每个区域都有重要作用。其中, Luminal结构域的低聚化被认为是启动信号在内质网膜上传播的关键事件, 它可以使IRE1的胞质激酶和RNase模块协同组装成一个有序的具有明确三维结构的低聚体[17]。在Ali等[18]的研究中发现, IRE1-like蛋白通过其N端内质网Luminal结构域的联合激活, 促进其胞质激酶结构域的自磷酸化, 从而激活C端的核糖核酸酶结构域, 该结构域可剪接Xbp1 mRNA, 生成一个活跃的Xbp1s转录激活因子。而STKc和RNase结构域是哺乳动物IRE1蛋白的主要效应区, 蛋白激酶活性结构域自身磷酸化, 将核糖核酸酶活性结构域激活并发挥其生理功能[8,19,20]。草鱼IRE1-like蛋白的三个保守结构域是否涉及这些生理功能有待研究。

图 3 MC-LR对草鱼肝脏IRE1-like 基因表达的影响Fig. 3 The effects of MC-LR on the expression of IRE1-like in liver of grass carp (P <0.05)
通过氨基酸同源性比对分析, 草鱼IRE1-like 氨基酸序列与已报道的物种的氨基酸具有较高的相似性和一致性, 其中与斑马鱼IRE1-α的相似性非常高, 达到了95%。而与人和小鼠IRE1-β的相似性较低, 且进化树分析显示草鱼IRE1-like与IRE1-β也没有聚为一支。已有的研究表明, 人和哺乳动物IRE1包括两种类型, 即IRE1-α和IRE1-β[8,9], 但在鱼类中没有IRE1-β的相关报道, 本研究的结果也表明草鱼IRE1-like不属于IRE1-β类型。因此, 从草鱼IRE1-like蛋白序列等分析来看, 草鱼IRE1-like与哺乳动物和其他鱼类的IRE1-α或IRE1-like类似, 这预示着IRE1-like在内质网应激中发挥着同样重要的作用。
3.2 IRE1-like基因在草鱼不同组织中的表达分析
研究表明,IRE1-α基因在人体内广泛表达, 而IRE1-β基因主要在胃、肠道和呼吸道上皮细胞中表达[2]。在水产动物中,IRE1基因的组织分布表达研究较少, 仅在黄颡鱼和南美白对虾中有报道[11,12]。IRE1-α基因在黄颡鱼广泛表达, 其中在肝脏中表达最高, 其次为心脏、脾脏、中肠、脑、头肾和精巢,在骨骼肌和鳃中的表达最低[11]。南美白对虾中IRE1基因在所检测的组织(鳃、胃、心脏、血液、表皮、肠道、幽门盲囊、肌肉、神经、肝胰腺和眼柄)中也呈现组成型表达, 其中在血液中表达最高[12]。本研究同样发现草鱼IRE1-like基因也可在所检测的不同组织中表达, 其中肝脏中的表达量相对最高, 这与IRE1-α基因在黄颡鱼中的表达比较相似[11]。研究基因的组织分布表达模式有助于了解该基因的生理功能,IRE1基因在水产动物各组织中的广泛表达说明它在水产动物中具有非常重要的作用, 可在不同组织中参与各种生理功能。
3.3 不同剂量MC-LR对草鱼肝脏IRE1-like基因的影响
IRE1是内质网膜上的感受蛋白, 被认为在内质网应激中起着非常重要的作用[11], 已有研究报道表明MC-LR可诱导细胞内质网应激[22—24], 而过度或长时间的内质网应激则会激活IRE1, 进而激活JNK,导致细胞凋亡[25]。UPR的激活可降低蛋白质的翻译, 从而增加蛋白质的折叠能力, 然而如果这种初始反应不能缓解内质网应激, 则UPR信号网络的长期激活会导致细胞凋亡[26]。本研究检测了不同剂量MC-LR诱导后草鱼肝脏中IRE1-like基因的表达,在25 μg/kg BW组诱导24h后, 表达显著升高, 而100 μg/kg BW剂量组草鱼IRE1-like基因的表达在诱导96h后显著升高。根据本实验已发表的文献可知草鱼注射25和100 μg/kg BW MC-LR 24h和96 h后均可诱导细胞凋亡, 尤其在100 μg/kg BW MCLR组中草鱼肝细胞凋亡更严重[16], 因此, 我们推测MC-LR可诱导内质网应激, 随后激活了草鱼IRE1-like基因并进一步导致了细胞凋亡。尽管25 μg/kg BW组草鱼肝脏IRE1-like基因在96h的表达相对于对照组表达升高了但差异不显著, 并呈现下降趋势,这可能与25 μg/kg BW MC-LR的剂量较低, 不会对内质网产生持续的应激有关, 也可能与IRE1自身能降解内质网中的错误蛋白, 从而释放内质网的负荷有关[7], 具体原因还有待于进一步深入研究。
综上所述, 本研究通过RACE技术克隆得到了草鱼IRE1-like基因cDNA的全长序列, 并对其编码的1030个氨基酸进行了一系列生物信息学分析, 研究了草鱼IRE1-like基因的组织表达分布模式及草鱼肝脏中IRE1-like基因对不同剂量MC-LR的响应,发现IRE1-like基因在不同组织的表达为组成型表达, 另外, MC-LR可促进草鱼肝脏中IRE1-like基因表达上调。后续有望通过RNAi等方法对草鱼IRE1-like基因的生物学功能包括在响应MC-LR中所起的作用进行更加深入的探索。
