达氏鲟Elovl4、ELovl5和Elovl7克隆、组织分布及饥饿对其表达的影响
2020-12-10吴晓雲陈叶雨赖见生宋明江
吴晓雲 陈叶雨 刘 亚 赖见生 宋明江 龚 全
(四川省农业科学院水产研究所, 成都 611731)
长链脂肪酸延长酶(Elongases of very long chain fatty acids, Elovls)位于细胞内质网上, 是长链多不饱和脂肪酸(Long chain polyunsaturated fatty, LCPUFA)合成的关键限速酶, 在维持细胞膜结构与功能、能量代谢和信号转导等生命活动中起重要作用[1,2]。目前在哺乳动物中已发现7种Elovls(Elovl1-Elovl7), 其中Elovl1、Elovl3、Elovl6和Elovl7主要参与长链饱和脂肪酸及单不饱和脂肪酸的合成, 而Elovl2、Elovl4和Elovl5主要参与合成多不饱和脂肪酸[3]。水生动物长链脂肪酸延长酶的研究相对较晚, 主要集中在Elovl2、Elovl4和Elovl5[2,4],Elovl7仅在榄绿青蟹中被克隆[5]。其中, Elovl4能将C20和C22的多不饱和脂肪酸最高延长到C36[4], 主要参与超长链脂肪酸的合成, 因而它主要在哺乳动物视网膜、脑和精巢等脂质含量较多的组织中表达[6]; 在哺乳动物中, Elovl5能够优先延伸C18和C20多不饱和脂肪酸, 而对C22多不饱和脂肪酸的活性较弱, 很难延长到C22以上[6,7], 鱼类的研究也保持一致[8,9];然而, 目前在鱼类中对Elovl7的研究较少, Elovl7可对C18和C16表现出较强的活性, 能将C18延长到C20再转移给Elovl1进一步延长到C24[10]。
达氏鲟(Acipenser dabryanus), 又称长江鲟, 沙腊子或小腊子。隶属于鲟形目(Acipenseriformes)、鲟科(Acipenseridae)、鲟属(Acipenser), 曾是长江上游重要的经济捕捞对象[11]。但由于水体污染、过度捕捞和栖息地的破坏, 达氏鲟微生境发生改变,极易因时间、空间、季节更替以及环境变化导致的食物短缺而造成饥饿胁迫。本研究通过克隆Elovl4、Elovl5和Elovl7得到了其cDNA开放阅读框序列, 并研究其在不同组织及饥饿胁迫作用下的表达模式, 以期为进一步了解饥饿胁迫条件下达氏鲟Elovl4、Elovl5和Elovl7表达变化情况, 为进一步探索达氏鲟生理生化变化机制及Elovl4、Elovl5和Elovl7功能奠定基础, 也为其资源的管理和人工养殖提供理论依据。
1 材料与方法
1.1 实验材料
实验用鱼为四川省农业科学院水产研究所人工繁殖的F2代达氏鲟(Acipenser dabryanus)幼鱼。实验前将鱼在循环水养殖系统暂养2周。每天早晚各投喂1次饲料, 饲料含量为粗蛋白48.62%、粗脂肪8.61%、粗纤维4.68%、粗灰分14.66%和水分12.21%。暂养2周后, 选取75尾体重为(30±2) g, 健康、活力好的达氏鲟随机分为5个处理, 每个处理3个平行, 每个平行5尾鱼, 分别为对照组(饥饿0)、实验1组(饥饿1d)、实验2组(饥饿3d)、实验3组(饥饿7d)和实验4组(饥饿14d)进行为期14d饥饿胁迫试验, 期间DO≥5.0 mg/L, 水温25—27℃。达氏鲟通过MS-222(Tricaine methanesulfonate)进行麻醉后,将不同组织(心、肝、脾、肾、卵巢、精巢、肌肉、肠、鳃、皮肤、脑和胃)取出后放入液氮中速冻, 然后置于–80℃冰箱保存备用。
1.2 RNA提取及cDNA合成
总RNA提取采用RNAisoPlus(宝生物工程大连有限公司, D9108B)按试剂盒说明书的方法操作。将–80℃冻结的组织样品在液氮中研磨至粉末状,加入1 mL RNAiso Plus, 在试剂的作用下从组织中抽提出总RNA, 溶解于50 μL DEPC水中。所得RNA溶液用1.0%琼脂糖凝胶电泳(120 V, 10min)检查其相对分子质量和完整性。RNA样品采用Nano Drop 1000分光光度计(Hach, America)检测其纯度和浓度。第一链cRNA合成参照ReverTra Ace反转录试剂盒(TOYOBO, Japan)进行操作。
1.3 达氏鲟Elovl4、Elovl5和Elovl7基因克隆
根据本实验室前期达氏鲟转录组测序获得的unigene序列为基础(Accession numbers: SRR616 7299、SRR6172670、SRR6173479、SRR617 5505、SRR6179331和SRR6179394), 通过NCBI数据库进行比对分析, 获得了达氏鲟Elovl4、Elovl5和Elovl7的cDNA全序列。为验证序列的正确性, 分别在ORF上下游设计验证引物(表 1), 以反转录cDNA为模板进行PCR扩增, 扩增产物连接载体后, 送生工生物工程(上海)股份有限公司进行测序。
1.4 序列分析
运用Vecter NTI软件分别预测达氏鲟Elovl4、Elovl5和Elovl7 ORF, 并推测其编码氨基酸序列, 计算编码产物分子量和等电点; 信号肽序列使用在线工具SignalP 4.0 (http://www.cbs.dtu.dk/services/SignalP/)进行预测; 跨膜结构分析用SMART(http://smart.embl-heidelberg.de/)。利用Cluxtal X将达氏鲟Elovl4、Elovl5和Elovl7氨基酸序列分别与其他物种的相应氨基酸序列进行多重比对。利用Mega 6.0软件的Neighbor-joining法构建系统进化树。

表 1 Elovl基因克隆和荧光定量PCR检测所用引物Tab. 1 Primers for cDNA cloning and real-time PCR of Elovls
1.5 组织表达分析
选取5尾健康达氏鲟, 采用荧光定量PCR法检测Elovl4、Elovl5和Elovl7在心、肝、脾、肾、卵巢、精巢、肌肉、肠、鳃、皮肤和脑中的相对表达量。荧光定量PCR扩增采用SYBR@PrimeSriptTMRT-PCR Kit Ⅱ(perfect Real Time, 宝生物工程大连有限公司, DRR083A)。10倍连续稀释的阳性模板作为PCR模板做标准曲线, 扩增效率在95%—105%,r2>0.9, 溶解曲线单一峰。结合琼脂糖电泳检测产物长度和特异性, 从而确定引物最优退火温度。12.5 μL 2×SYBR Green Real-time PCR Master Mix,上下游引物各1 μL(10 μmol/L), 灭菌双蒸水5.5 μL。PCR扩增程序为: 94℃预变性5min; 94℃变性30s,55℃退火30s, 72℃延伸30s, 42个循环; 最后70—95℃获取熔解曲线并确保扩增产物的特异性。反应完成后检测产物的特异性。每个样品设3个复孔。采用2–ΔΔCt法分别计算样品中Elovl4、Elovl5和Elovl7 mRNA的相对表达量。
1.6 饥饿胁迫下达氏鲟Elovl4、Elovl5和Elovl7表达量的检测
实验鱼分别在饥饿实验的0、1d、3d、7d和14d采集肌肉、脑、胃、肠道及肝脏组织, 每个平行采集3尾。荧光定量方式参考1.5。
1.7 数据分析
采用平均值±标准误(Mean±SE)表示实验数据,采用SPSS22.0软件处理结果。对试验数据进行方差分析并结合Duncan法进行多重比较检测显著性。
2 结果
2.1 达氏鲟Elovl4、Elovl5和Elovl7克隆及序列分析
达氏鲟Elovl4编码区为753 bp, 编码250个氨基酸, 相对分子质量为29793.13 Da, 等电点为9.47, 包含7个预测的跨膜区;Elovl5编码区为885 bp, 编码294个氨基酸序列, 相对分子质量为34794.38 Da, 等电点为9.42, 5个预测的跨膜区;Elovl7编码区为846 bp, 编码281个氨基酸序列, 相对分子质量为33227.82 Da, 等电点为9.19, 7个预测的跨膜区。Elovl4,Elovl5和Elovl7均含还有高度保守的HXXHH组氨酸盒。
2.2 进化树分析
将达氏鲟Elovl4、Elovl5和Elovl7氨基酸序列分别与其他鱼类相应序列进行比对后发现, 三个基因均与斑点雀鳝聚为一支, 表明在进化上与雀鳝亲缘较近(图 1)。
2.3 达氏鲟不同组织Elovl4、Elovl5和Elovl7的表达
达氏鲟Elovl4、Elovl5和Elovl7在各个组织中均有表达, 同时存在着差异性表达(图 2),Elovl4在卵巢和眼中表达最高, 其次是脑(P<0.05), 其他组间没有显著差异(P>0.05);Elovl5在肌肉中表达最多,其次是眼(P<0.05), 其他组间没有显著性差异(P>0.05);Elovl7在卵巢中表达最高(P<0.05), 鳃、精巢和肌肉中表达较多(P<0.05), 其他组间没有显著差异(P>0.05)。
2.4 饥饿胁迫下达氏鲟Elovl4、Elovl5和Elovl7表达量的变化
在饥饿胁迫下, 达氏鲟Elovls在不同组织中的表达各有不同。饥饿7d时(图 3A),Elovl4在脑、肠道和肌肉中的表达量显著提高(P<0.05), 之后显著下降(P<0.05); 饥饿0和1dElovl4在胃中的表达没有显著差异(P>0.05), 之后显著下降, 14d时显著上调(P<0.05); 肝脏中Elovl4在0时表达量最高, 随后显著下降(P<0.05), 其他组间没有显著性差异(P>0.05)。
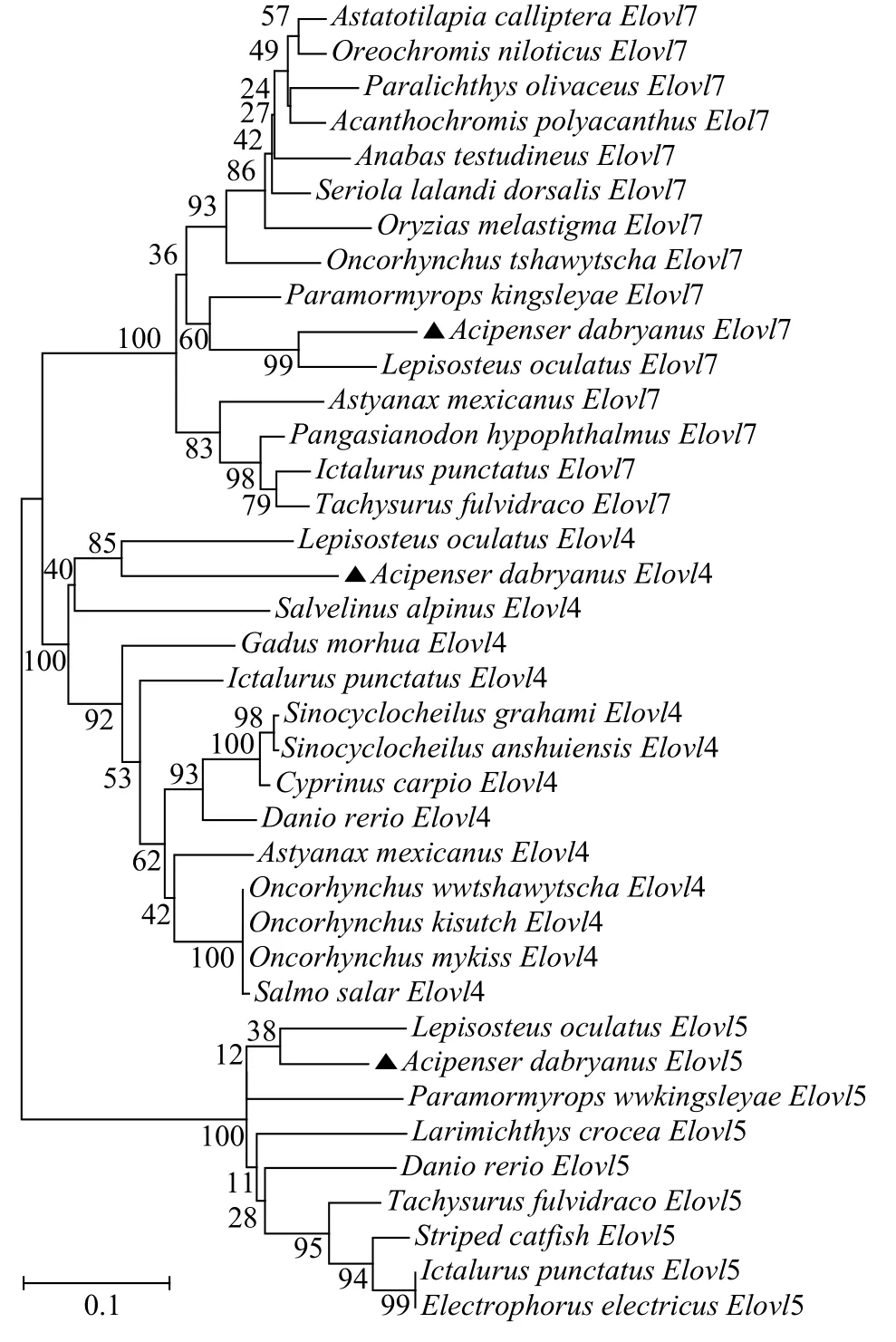
图 1 达氏鲟Elovl4、Elovl5和Elovl7与其他脊椎动物的系统发生树Fig. 1 Phylogenetic tree of the Elovl4, Elovl5 and Elovl7 from other vertebrates based on neighbor-joining method
饥饿7d时(图 3B),Elovl5在脑和肌肉中的表达量显著提高(P<0.05), 之后显著下降(P<0.05); 饥饿3d时Elovl5在胃中的表达量最低, 且显著低于1d、7d和14d组; 肝脏中Elovl5表达量随时间推移逐渐下降, 0时表达量显著高于其他各组(P<0.05), 1d、3d和7d组间没有显著性差异(P>0.05), 14d表达量最低; 肠道中Elovl5表达量逐渐升高, 14d时显著高于其他各组(P<0.05)。
Elovl7在脑和肌肉中的表达量先下降后在7d时显著提高(图 3C), 随后显著下调(P<0.05); 3d组Elovl7在胃中的表达量最低, 显著低于0和14d组, 与1d和7d组间没有显著性差异(P>0.05),Elovl7在胃中的表达量在0时最高, 之后显著下降(P<0.05), 其余各组间没有显著性差异(P>0.05); 肝脏中Elovl7表达量随时间推移逐渐下降, 0时表达量显著高于其他各组(P<0.05); 肠道中Elovl7表达量在0和14d时显著高于其他各组(P<0.05), 其他组间没有显著性差异(P>0.05)。
3 讨论
脂肪酸是鱼类生长所必须的营养物质, 同时还具有重要的生理学功能。长链多不饱和脂肪酸(LC-PUFA), 主要包括二十碳五烯酸(EPA, 20:5n-3)、二十二碳六烯酸(DHA, 22:6n-3)和花生四烯酸(ARA, 20:4n-6)等, 对细胞膜的完整性和流动性、转录调控以及细胞信号转导有重要作用[12]。Elovls作为合成长链脂肪酸的关键限速酶, 其重要性显而易见。
3.1 达氏鲟Elovl4、Elovl5和Elovl7的组织表达差异
Elovl4在斑马鱼(Brachydanio rerio)[13]、军曹鱼(Rachycentron canadum)[14]、大西洋鲑(Salmo salar)[4]、河豚(Tetraodontidae)[15]和大黄鱼(Larimichthys crocea)[16]等中都得到克隆, 斑马鱼中存在2个亚型,Elovl4a和Elovl4b, 分别编码309和303个氨基酸[13], 西洋鲑[4]Elovl4编码306个氨基酸, 而本研究结果显示, 达氏鲟Elovl4仅编码250个氨基酸, 说明在进化过程中鲟Elovl4 CDS区能编码氨基酸数与其他鱼类有较大的差异。Elovl4主要在哺乳动物视网膜、脑和精巢等脂质含量较多的组织中表达[6]。鱼类与哺乳动物类似, 在大西洋鲑[4]的研究中发现,Elovl4主要在眼中表达; 军曹鱼[14]Elovl4主要在脑、眼和垂体中表达;Elovl4在斑马鱼上的2个亚型,Elovl4b主要在眼、卵巢和精巢中表达, 而Elovl4a除了在肌肉和脂肪组织中没有检测到表达信号以外,在其他组织中均有表达; 河豚[15]眼和脑中均能检测到信号。结合本实验结果发现, 达氏鲟Elovl4除了在眼中表达较多以外还在脑和卵巢中有较多的表达, 与前人的研究基本保持一致, 推测Elovl4与达氏鲟视觉发育和繁殖功能密切相关。
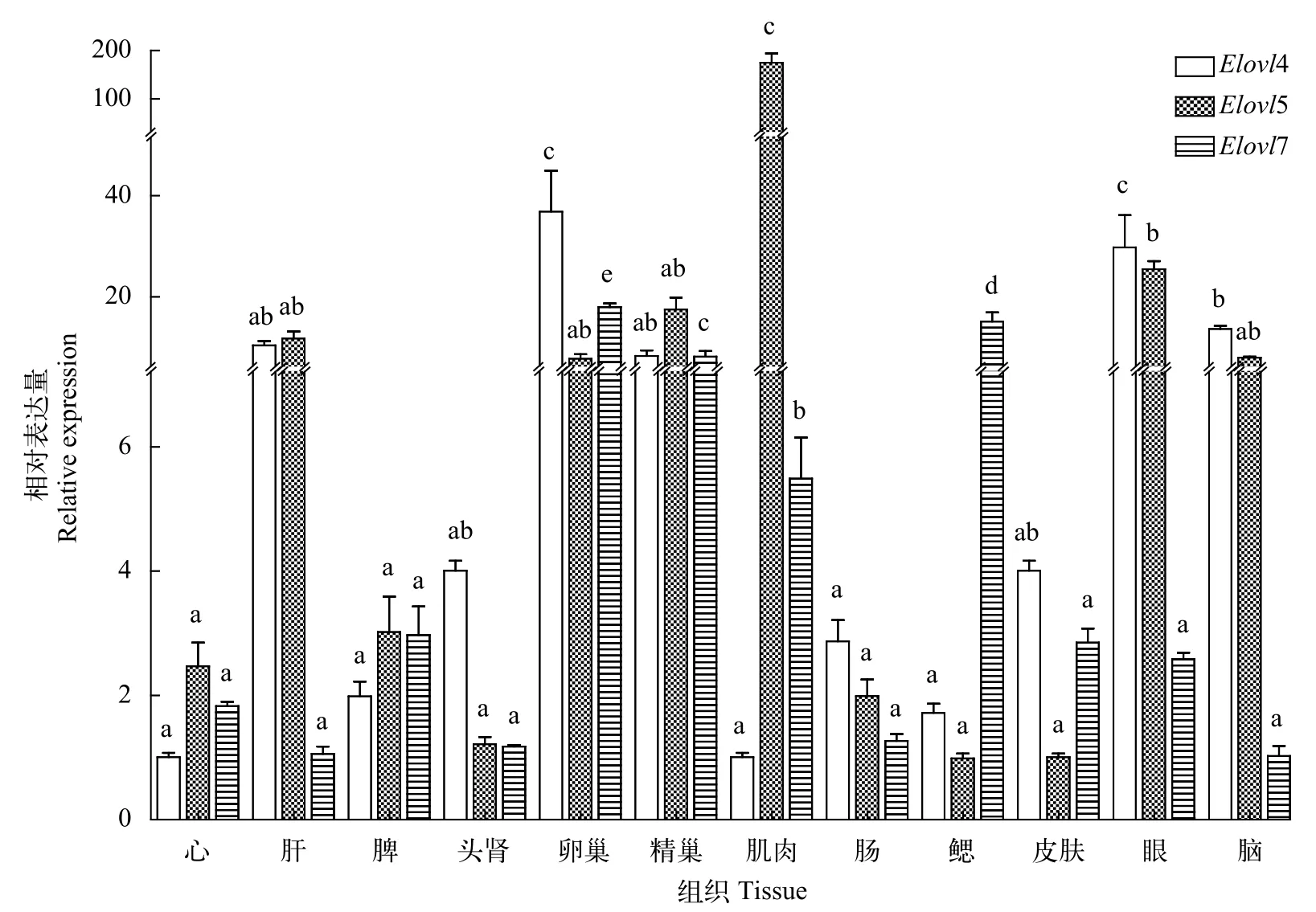
图 2 不同组织中Elovl4、Elovl5和Elovl7 mRNA的相对表达量Fig. 2 The relative expression of Elovl4, Elovl5 and Elovl7 mRNA in different tissues
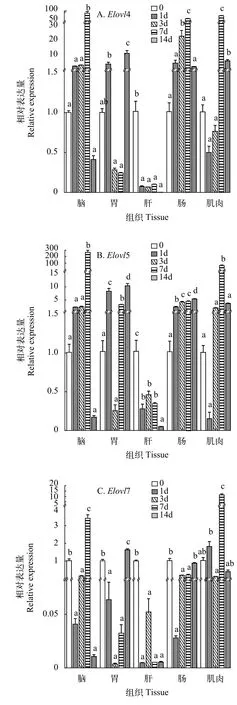
图 3 饥饿胁迫下不同组织中Elovl4、Elovl5和Elovl7 mRNA的相对表达量Fig. 3 Effect of starvation on the relative expression of Elovl4,Elovl5 and Elovl7 mRNA in different tissues (n=9)
鱼类Elovl5最先是在斑马鱼[17]上克隆得到, 可编码氨基酸291个, 研究相继发现河豚[15]、镜鲤(Cyprinus carpio)[18]和银无须魮(Puntius gonionotus)[19]Elovl5编码291个, 双棘黄姑鱼(Nibea diacanthus)[20]、南方黑鲔(Thunnus maccoyii)[21]、日本鳗鲡(Anguilla japonica)[8]和军曹鱼[22]Elovl5编码294个, 大西洋鲑[23]Elovl5有两个亚型,Elovl5a编码295个,Elovl5b编码294个。本实验达氏鲟Elovl5序列预测跨膜区为5个, 与军曹鱼[22]、河豚[15]和大西洋鲑[23]Elovl5b结果类似。不同种类的鱼类Elovl5在组织中的表达存在差异, 但主要还是在肠道、肝脏和脑中表达。塞内加尔鳎(Solea senegalensis)[24]Elovl5在肝脏和肠道中表达较多, 而脑次之, 鳃、心脏、中肾和肌肉中未检测出信号; 河豚[15]Elovl5主要在肝脏、脑、肠道和眼中表达; 军曹鱼[22]Elovl5在脑、肝脏和心脏中表达较多, 皮肤和鳃中没有检测到信号; 银无须魮[19]Elovl5主要在肝脏和肠道中表达, 其次为脑和肌肉; 日本鳗鲡[8]Elovl5主要在脑、肝脏和肠道中表达较多; 双棘黄姑鱼[20]和大西洋鲑[23]主要在肠道和肝脏中表达。达氏鲟Elovl5在肌肉和眼中表达较多, 脑和肝脏次之, 与其他鱼类上的研究基本类似, 可能是由于实验所用达氏鲟正处于生长发育重要时期, 其肌肉中脂肪代谢活动旺盛,这与本实验室之前对脂肪酸结合蛋白的研究结果相一致(数据未发表)。另外, 这也暗示Elovl5对达氏鲟视觉的发育有密切联系。
目前对Elovl7的研究较少, 研究发现橙泥蟹(Scylla olivacea)[25]Elovl7编码350个氨基酸, 苏太猪(Sus)[26]Elovl7编码281个氨基酸, 鸡(Gallus gallus)[27]Elovl7编码279个氨基酸。牛(Bovine)Elovl7在肾脏中表达较多, 在肌肉、胃和肝脏中较少[28],Elovl7在30周龄鸡十二指肠、腺胃、肾脏和肺中表达较多[27]; 橙泥蟹Elovl7, 在胃中表达最高, 其次是肠道和鳃[25]。本实验结果显示, 达氏鲟Elovl7主要在卵巢、鳃、精巢和肌肉中表达, 这与哺乳动物还是存在差异的, 但与橙泥蟹类似的是在鳃中的表达相对较高, 说明Elovl7在水生动物的鳃中发挥了一定作用, 但具体原因还有待研究。水生动物中对Elovl7的报道较少, 在哺乳动物上已有证明Elovl7与前列腺肿瘤细胞的生长有关[27], 在仅有的研究中发现它可能在脂肪细胞分化的过程中出发脂质积累[30], 而Elovl7在达氏鲟脂肪代谢中的作用和功能还有待进一步研究。
3.2 饥饿对达氏鲟Elovl4、Elovl5和Elovl7表达差异的影响
一般来说, 鱼类自身不能或仅少量合成n-3和n-6系列不饱和脂肪酸, 主要通过外界摄取获得, 淡水鱼类的必需脂肪酸主要包括亚油酸(18﹕2n-6)、α-亚麻酸(18﹕3n-3)、γ-亚麻酸(18﹕3n-6)[31]。禁食可以提高肝脏的分解代谢率, 反映了生存所需储备的调动[32]。肝脏是脂肪酸合成的重要部位[33], 在禁食的条件下, 不饱和脂肪酸摄入减少, Elovls底物减少,导致Elovls合成下降, 这或许可以解释肝脏中的Elovl4、Elovl5和Elovl7转录水平随时间的增加直线下降的现象。
脑是调节动物生长和发育的重要中枢系统, 肌肉反映生长状况的重要组织, 而鱼类的脑和肌肉中含有丰富的不饱和脂肪酸[34]。禁食过程中的鱼类对脂肪酸的代谢, 一般先从饱和脂肪酸开始, 其次是低不饱和脂肪酸, 最后才动用高不饱和脂肪酸[35,36]。这或许是达氏鲟饥饿第7天时脑和肌肉组织中Elovl4、Elovl5和Elovl7的转录水平显著提高的原因之一, 由于饥饿后期高不饱和脂肪酸水平下降,Elovl4、Elovl5和Elovl7转录水平上调, 之后可能由于底物减少使其转录水平下降; 这也暗示脑与肌肉中的脂质代谢不同于肝脏。另外, 黄红丽等[37]对大黄鱼的研究发现, 饥饿使大黄鱼肌肉组织中I-FABPb表达量先上调后下降, 这与本实验中的结果相一致。
长期的饥饿能显著影响鱼类消化器官的组织结构[36], 其结构的改变必然引起功能的障碍。胃肠道是食物消化和吸收的重要场所, 尤其是肠道, 承载着消化和吸收双重功能, 并通过肠道绒毛结构增大吸收面积, 其上皮细胞的形成及其流动性都与不饱和脂肪酸密切相关。本实验的结果显示, 达氏鲟胃Elovl4、Elovl5和Elovl7三个基因在饥饿3d时均有显著的下调, 表明饥饿对达氏鲟胃组织LC-PUFA合成可能存在抑制作用。另外, 我们还发现, 在饥饿条件下Elovl4、Elovl5和Elovl7在达氏鲟肠道中的表达趋势完全不同, 在饥饿1d、3d和7d时Elovl4和Elovl5 mRNA水平于对照组相比呈现增加的趋势, 而Elovl7 mRNA水平却显著下降, 在一定程度上也表明了脂质代谢平衡调节过程复杂[38], 3个基因在肠道中有不同的功能, 具体原因有待进一步研究。
综上所述, 本研究首次克隆了达氏鲟Elovl4、Elovl5和Elovl7 CDS序列, 并分析了其组织分布和在饥饿胁迫下的表达模式, 发现Elovl4、Elovl5和Elovl7在达氏鲟视觉、脑、性腺以及肌肉的发育与生长过程中可能发挥着重要的作用。研究还发现饥饿胁迫对不同组织中的Elovls作用不同, 能显著影响达氏鲟Elovl4、Elovl5和Elovl7 mRNA的转录水平, 产生差异的原因可能与其主要分布部位以及所参与合成的LC-PUFA功能有关, 具体原因还有待研究, 但能肯定的是饥饿应激能显著影响肝脏的脂质代谢, 并且可能通过降低Elovl4、Elovl5和Elovl7转录来抑制LC-PUFA合成。
