NLRP3介导线粒体损伤在非酒精性脂肪性肝炎中的作用及SS-31的干预研究*
2020-11-26林晔
林晔
非酒精性脂肪性肝炎(Nonalcoholic steatohepatitis,NASH)是非酒精性脂肪性肝病(Nonalcoholic fatty liver disease,NAFLD)的主要进展形式,具有较高的肝纤维化和肝癌风险,主要病理特征包括脂肪变性、气球样变、肝细胞损伤、混合性炎细胞浸润和不同程度的纤维化[1-2]。近期研究发现,线粒体结构和功能异常参与NASH 的起病与进展,炎症小体在炎症以及纤维化进展中起到关键作用[3-6]。本研究旨在明确NLRP3 介导线粒体损伤在NASH 中的作用机制,以及具有线粒体保护作用的线粒体靶向抗氧化肽SS-31 对NLRP3 信号通路的作用以及对NASH 的影响,以寻求干预NASH的有效药,现报道如下。
1 材料与方法
1.1 材料与试剂 选择健康8 周龄雄性C57BL/6J 小鼠共30 只,体重150~180 g,平均(171.33±10.12)g,购自中国科学院动物试验中心。SS-31(广州天源生物科技有限公司)、HE 染液(上海莼试生物技术有限公司)、Caspase-1 一抗、IL-1β 一抗、IL-18一抗、Caspase-1 二抗、IL-1β 二抗、IL-18 二抗(均购自美国Abcam 公司)、DAB 显色液(广州鼎国生物技术有限公司)、苏木素(成都格利普生物科技有限公司)、Trizol 试剂(深圳时代生物科技有限公司)。
1.2 方法
1.2.1 构建NASH 模型 采用胆碱-蛋氨酸缺乏(MCD)饲料喂养法,连续喂养8 周,脱颈法处死获得肝脏组织,HE 染色病理观察肝脏出现脂肪变、炎症浸润和纤维化,符合NASH 病理改变为造模成功。
1.2.2 实验分组和观察指标 30 只小鼠按随机数字表法分为对照组、模型组和干预组,每组10 只。对照组正常饮食喂养,模型组和干预组采用MCD 饲料喂养。干预组腹腔注射SS-31 3 mg/kg,1 次/周;对照组注射等量0.9%氯化钠溶液,共持续喂养8 周。观察各组小鼠体重,使用生化分析仪检测血清丙氨酸氨基转移酶(ALT)和天门冬氨酸氨基转移酶(AST)含量;HE 染色观察肝脏组织病理变化;实时定量PCR(RT-PCR)法检测肝脏线粒体损伤指标,包括ATPase-6 和细胞色素C 基因表达量;改良氧电极法测定线粒体呼吸控制率(RCR)与线粒体琥珀酸脱氢酶(SDH)活性;免疫组化染色检测肝脏NLRP3 表达量;Western blot 法检测肝脏Caspase-1、白介素(IL-1β)和IL-18 表达水平。
1.2.3 免疫组化染色 肝脏组织石蜡切片厚度4 μm,经脱蜡、水化、抗原修复后滴加兔抗鼠NLRP3 抗体(工作浓度1︰2 000)置于湿盒内4 ℃孵育过夜,以正常小鼠IgG 作为阴性对照;PBS 洗涤后滴加羊抗兔抗体(工作浓度1︰500)置于湿盒中27 ℃孵育20 min;PBS 洗涤后滴加辣根酶标记链霉卵白素工作液置于湿盒中27 ℃孵育20 min;PBS 洗涤、DAB 显色、苏木素复染等操作,于光学显微镜下观察,采用半定量法,计算黄染阳性细胞百分比。
1.2.4 RT-PCR 法 Trizol试剂提取细胞中总RNA,测定浓度后根据反转录试剂盒提示合成cDNA,设计引物序列,ATPase-6:上游5’-GGTTTCATCCAGGATCGAGCAGG-3’,下 游5’-ACAAAGATGGTCACGGTCTGCC-3’;细胞色素C:上游5’-ACTACTTCTCCCGCCGCTAC-3’,下游5’-GAAATCAAACAGAGGCCGCATG-3’;内参U6:上游5’-CGCGAGAAGATGACCCAGAT-3’,下游:5’-GCACTGTGTTGGCGTACAGG-3’。根据反应试剂盒提示配置反应体系和设置反应参数,ATPase-6 和细胞色素C 基因表达量结果以2-⊿⊿Ct法表示。
1.2.5 Western blot 法 组织匀浆后加RIPA 裂解液提取细胞总蛋白,测定蛋白浓度后使用β-actin 抗体进行剂量标准化;取30 μg 样品蛋白和内参蛋白,经8% SDS-PAGE 电泳分离,将分离区带电转移至PVDF 膜,滴加羊抗鼠Caspase-1、IL-1β 和IL-18抗体(1︰2 000)静置过夜,PBS 洗涤后滴加兔抗羊对应抗体(1︰500)室温孵育4 h,PBS 洗涤,ECL显色。结果行半定量分析,以样品蛋白与内参蛋白的电泳条带灰度值的比值表示。
1.3 统计学处理 采用SPSS 20.0 软件对所得数据进行统计分析,计量资料用()表示,三组间比较采用单因素ANOVA 分析,组间比较采用独立样本t 检验。以P<0.05 为差异有统计学意义。
2 结果
2.1 三组喂养前、喂养2、4、8 周时体重和血清AST 和ALT 含量比较 三组小鼠均成功喂养8 周,无死亡。喂养前,三组体重、血清AST 及ALT 的含量比较,差异均无统计学意义(P>0.05)。喂养2~8 周,对照组体重、血清AST 和ALT 含量的波动幅度不显著,差异均无统计学意义(P>0.05);喂养2~8 周,模型组小鼠体重逐渐减小,血清AST 和ALT 含量均升高,差异均有统计学意义(P<0.05)。喂养2、4、8 周,干预组均体重高于模型组,而血清AST 和ALT 含量均低于模型组,差异均有统计学意义(P<0.05)。见表1 和图1。
表1 三组喂养前、喂养2、4、8周时体重和血清AST和ALT含量比较()
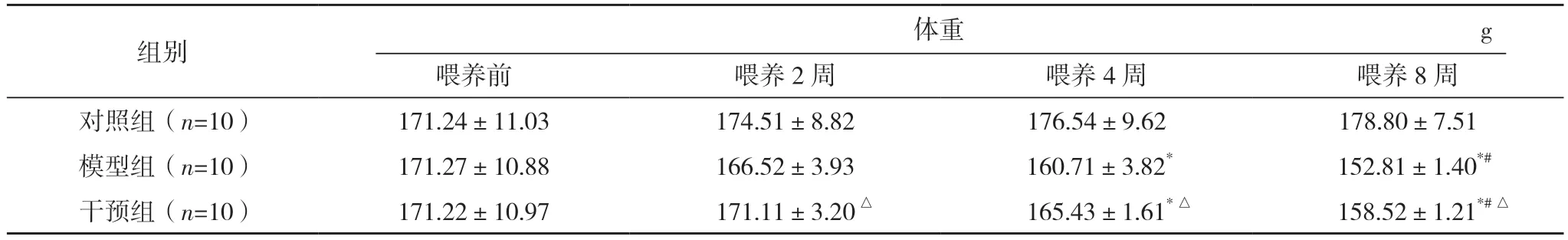
表1 三组喂养前、喂养2、4、8周时体重和血清AST和ALT含量比较()

表1 (续)
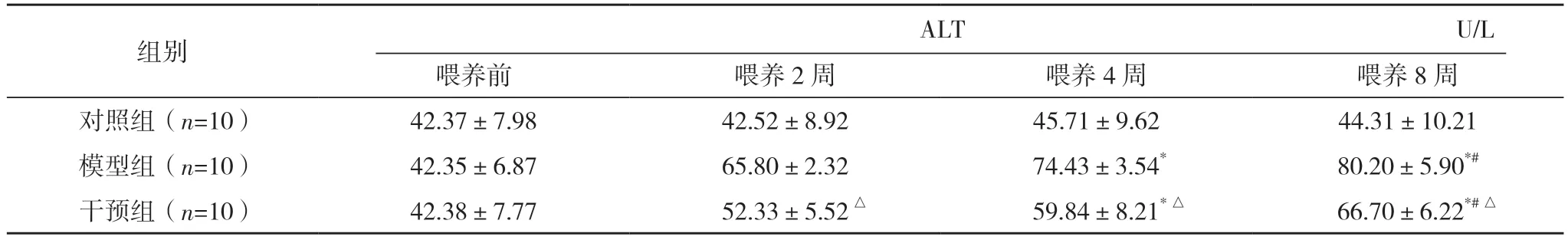
表1 (续)

图1 三组喂养2、4、8周时体重、血清AST和ALT含量的比较
2.2 HE 染色观察喂养8 周三组肝脏组织变化对照组肝脏无脂肪变,模型组出现不同程度的脂肪变、炎症浸润和纤维化,符合NASH 病理改变,造模成功;干预组上述变化较模型组减少,见图2。
2.3 三组ATPase-6 和细胞色素C 基因表达情况比较 模型组ATPase-6 和细胞色素C 基因表达量均明显高于对照组,差异均有统计学意义(P<0.05)。干预组ATPase-6 和细胞色素C 基因表达量均低于模型组,差异均有统计学意义(P<0.05),见表2与图3。
表2 三组ATPase-6 和细胞色素C基因表达()
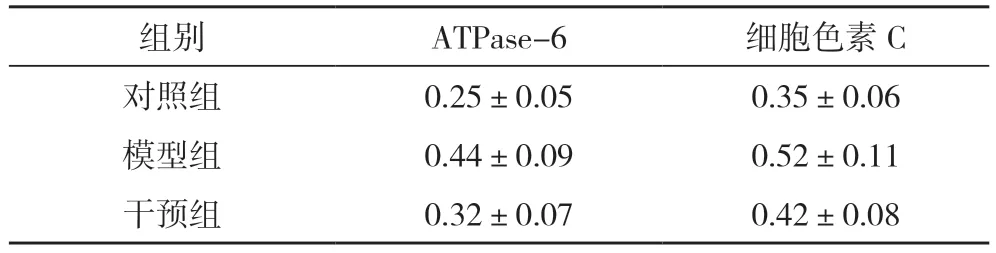
表2 三组ATPase-6 和细胞色素C基因表达()
2.4 三组线粒体RCR 和SDH 活性比较 模型组线粒体RCR 高于对照组,而SDH 活性低于对照组,差异均有统计学意义(P<0.05)。干预组线粒体RCR 低于模型组,而SDH 活性高于模型组,差异均有统计学意义(P<0.05)。见表3 和图4。
表3 随着线粒体RCR和SDH活性比较()

表3 随着线粒体RCR和SDH活性比较()

图2 HE染色观察喂养8周三组肝脏组织变化(100×)

图3 RT-PCR法检测三组ATPase-6 和细胞色素C基因表达量

图4 三组肝脏线粒体RCR和SDH活性比值
2.5 三组NLRP3 表达量比较 模型组NLRP3 阳性表达量(35.61±9.40)%高于对照组(6.92±2.32)%,差异有统计学意义(P<0.05)。干预组NLRP3 阳性表达量(16.72±5.51)%低于模型组,差异有统计学意义(P<0.05)。见图5、6。

图5 免疫组化染色检测三组NLRP3阳性表达(400×,箭头指NLRP3)
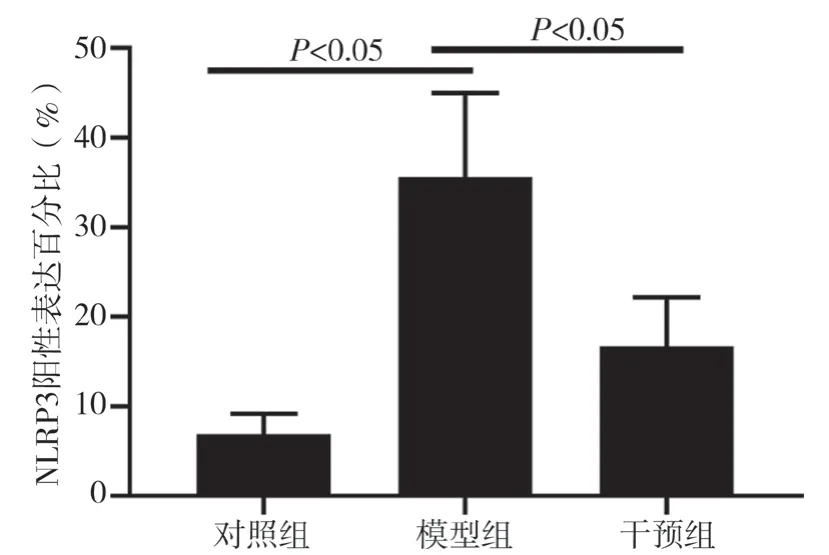
图6 三组NLRP3表达量比较
2.6 三组Caspase-1、IL-1β 和IL-18 表达比较 模型组Caspase-1、IL-1β 和IL-18 表达水平均高于对照组,差异均有统计学意义(P<0.05)。干预组Caspase-1、IL-1β 和IL-18 表达水平均低于模型组,差异均有统计学意义(P<0.05)。见表4 和图7。
表4 三组Caspase-1、IL-1β和IL-18表达水平比较()

表4 三组Caspase-1、IL-1β和IL-18表达水平比较()

图7 Western blot法检测三组肝脏组织Caspase-1、IL-1β和IL-18表达水平
3 讨论
研究发现,非甘油脂肪酸代谢产物在脂毒性和NASH 的发生发展中起重要作用,代谢相关氧化应激、自噬、炎症被认为是NASH 进展的重要标志[7-8]。通过本研究发现,随着喂养时间延长,模型组体重逐渐减小,血清AST 和ALT 含量升高,同时间点比较,干预组体重高于模型组,而血清AST 和ALT含量低于模型组,干预组ATPase-6、细胞色素C基因表达量、线粒体RCR 与NLRP3 阳性表达量均低于模型组,而SDH 活性高于模型组(P<0.05);干预组Caspase-1、IL-1β 和IL-18 表达水平均低于模型组(P<0.05)。
NAFLD 时线粒体功能降低,ATP 合成减少,ROS 消耗量减少但含量仍在增加,可能引起肝脏脂质沉积,从而加剧肝脏的氧化应激反应,导致肝细胞凋亡[9-10]。具体表现:线粒体脂肪酸氧化为机体提供能量,氧化异常导致游离脂肪酸进入肝脏含量增加,诱导氧化应激反应,损伤线粒体功能,氧化应激加剧肝细胞损伤和凋亡,氧化应激作为重要病理因素,是启动和维持NAFLD 的关键[11-13]。
炎症是NASH 发生以及纤维化进展的重要致病因素,NASH 患者体内存在促炎和抗炎细胞因子的失衡,其中NLRP3 炎性小体激活在NASH 进展中发挥重要作用。NLRP3 炎症小体活化机制主要有胞内离子浓度、线粒体功能异常和溶酶体损伤三种,NLRP3 炎症小体活化后招募并激活Caspase-1,Caspase-1 是NLRP3 炎性小体的效应蛋白,可将无活性的促炎细胞因子IL-1β 和IL-18 前体剪切为成熟的IL-1 和IL-18,从而引发机体炎症反应[14-16]。NLRP3 炎症小体信号通路可促进机体由炎症状态向纤维化进展。炎症刺激下肝星状细胞及肝细胞NLRP3 炎症小体表达明显升高[17]。肝损伤后IL-1表达增多,IL-1 受体缺陷可以缓解肝损伤及肝纤维化。提示IL-1 可能参与了从肝损伤到肝纤维化发生的整个过程[18]。Casepase-1 表达缺失可以缓解酒精性肝纤维化和非酒精性肝纤维化,提示Casepase-1同样参与肝纤维化的发生发展。临床研究表明,肝硬化患者血清IL-18 水平明显高于对照组,血清IL-18 水平与肝硬化患者Child-Pugh 评分及组织学损伤呈现正相关[19]。SS-31 是一种能够渗入细胞并且在线粒体内膜靶向定位聚集的小分子肽,SS-31在多种疾病中具有良好的抗氧化及抗凋亡作用[20]。
综上所述,NLRP3 炎性小体介导线粒体损伤参与了NASH 的发病过程,线粒体靶向抗氧化肽SS-31 可降低NASH 肝脏NLRP3 表达,改善线粒体损伤程度。
