GmXTH23基因的克隆及抗旱性鉴定
2020-11-18张沿政李永光李文滨
陈 龙,张沿政,李永光,李文滨
(东北农业大学农学院/大豆生物学教育部重点实验室/农业部东北大豆生物学与遗传育种重点实验室,黑龙江哈尔滨 150030)
【研究意义】据相关研究[1],我国耕地面积中干旱与半旱地区约占45%。干旱灾害极大地限制了作物产量,使粮食严重减产。大豆的产量与品质受干旱影响严重,因此,发掘优质的抗旱基因,是提高大豆抗旱性的有效途径。【前人研究进展】木葡聚糖水解酶/内转糖苷酶(XTH)是重构细胞壁的重要物质,其机制是通过催化作用,使木葡聚糖分子断裂重连,对植物的纤维素-木葡聚糖结构进行修饰从而影响细胞壁结构[2]。按照序列特征,XTHs被划分为Ⅰ、Ⅱ和Ⅲ类[3-4],其中III类被划分为IIIA和IIIB两个小类[5]。IIIB类、I类和II类的XTHs能内切木葡聚糖分子,产生还原性的末端,再和另一个木葡聚糖链相连接,其木葡聚糖内转糖苷酶(XET)活性非常显著[5-7]。IIIA类则显示出木葡聚糖水解酶(XEH)活性,专一水解木葡聚糖的β-1,4糖苷键。XTH酶的特征序列是DEIDFEFLG,此序列中包含了能够介导催化活性的氨基酸残基。该催化位点附近的苏氨酸或丝氨酸残基由N-糖基化修饰[7-8],与酶活性有重大关联。N-糖基化位点保守在IIIA亚类中未被发现[5],但在XTHs蛋白I/II类中已经被发现。XTHs蛋白的C-末端可以形成能够稳定蛋白质结构的二硫键,因为其通常含有高度保守的半胱氨酸。前人的研究成果[9]表明XTH与植物组织的的生长发育相关,如在拟南芥中AtXTH31参与了根的生长伸长;BcXTH1的过表达促进了花枝的生长从而使得株高增加[10];AtXTH15、19、16、17能够通过调节细胞壁延展性来促进叶柄伸长[11];在梨果成熟软化过程中PC-XET1可能参与了细胞壁的降解[12]。近期有研究[13-15]表明XTH还受环境胁迫影响,如在重力作用下拟南芥中AtXTH22表达量明显上升;在缺氧条件下,矮慈姑中的一些XTH基因表达上调[16];在拟南芥中xth15突变体与野生型相比,植株的耐铝性提高[17];过表达CaXTH3能够增强转基因拟南芥的抗旱性和耐盐性[18]。
【本研究切入点】通过本课题组前期研究发现,大豆基因GmXTH23在干旱胁迫下表达量显著增加,但是在植物抗旱生理中行使的功能尚未明确。【拟解决的关键问题】本试验通过同源克隆的方法从垦丰16中得到基因GmXTH23;利用qRT-PCR技术对基因在大豆不同组织内和模拟干旱胁迫下的表达情况进行分析;通过检验目的基因与黄色荧光蛋白同源重组质粒的蛋白质产物进行基因亚细胞水平定位;利用蘸花法侵染拟南芥得到过表达株系,同时对过表达株系进行干旱胁迫,观察表型并测量相关生理指标,验证GmXTH23基因在植物抗旱生理中的作用,探究其作用机制,为进一步研究GmXTH23基因功能提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 植物材料 本研究所用大豆材料垦丰16,拟南芥材料是由实验室自行繁殖并保存的哥伦比亚野生型(Columbia-0)。
1.1.2 主要试剂 反转录试剂盒、胶回收试剂盒、qRT-PCR 试剂盒、限制性内切酶、T4DNA 连接酶均于Takara公司购置。农杆菌感受态GV3101,大肠杆菌感受态DH5α均由实验室保存。
1.2 方法
1.2.1 大豆基因GmXTH23的克隆 采用Trizol 试剂提取大豆品种垦丰16 的幼叶的总RNA,用Takara 反转录试剂盒将其反转录成cDNA。利用Prime5.0 设计特异性引物GmXTH23(表1)进行扩增。PCR 产物回收纯化后,采用T4连接酶连接至线性化的pCXSN(XcmI单酶切)载体并进行测序。
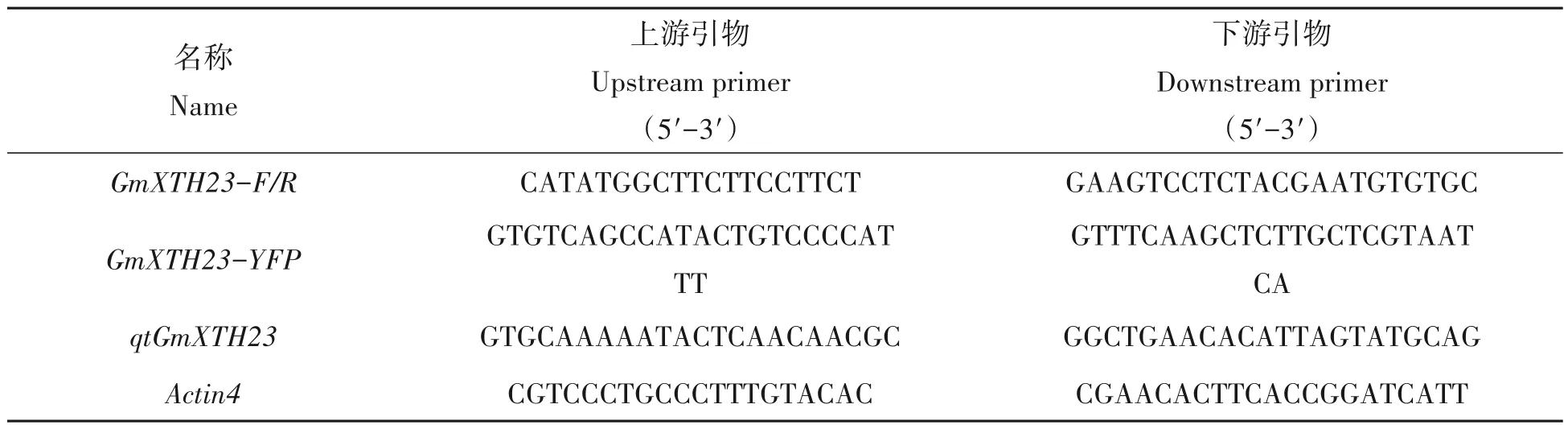
表1 引物序列Tab.1 Primer sequence
1.2.2GmXTH23基因的组织表达及胁迫下的表达模式分析 将垦丰16大豆培养于长日照(16 h光照/8 h黑暗),25 ℃条件下,当幼苗第2片三出复叶完全展开时,将幼苗挖出,水培24 h后分成两份,取一份置入含有5%PEG6000的水培液中,一份置于正常水培液中,分别取处理0,2,4,6,8,12,24,48 h的植物叶片,液氮迅速冷冻后保存于-80 ℃冰箱。提取以上样本的总RNA 将其反转录为cDNA,并将其作为模板,根据GmXTH23的序列信息设计定量引物qtGmXTH23(表1)进行qRT-PCR 分析,内参基因选用大豆持家基因Actin4。
1.2.3 GmXTH23 蛋白亚细胞定位 以1.2.1 中得到的GmXTH23胶回收产物为模板,以GmXTH23-YFP(表1)为引物扩增得目的片段,与YFP载体相连,测序比对。利用基因枪法将重组质粒在洋葱表皮细胞内进行瞬时表达,为了更准确地得到GmXTH23蛋白的定位信息,采用蔗糖溶液对细胞进行质壁分离,再次观察GmXTH23-YFP的表达情况。
1.2.4GmXTH23的遗传转化 将1.2.1 中得到的过表达载体pCXSN-GmXTH23采用冻融法将其转入农杆菌感受态GV3101 中。利用蘸花法侵染野生型拟南芥,得到T0 代过表达拟南芥株系并将其种子在含有草铵膦的MS培养基上进行筛选,繁殖得到T3代基因纯合的过表达株系用于后续试验。
1.2.5 表型试验 将野生型与T3 代纯合的过表达株系种子经10%次氯酸钠溶液消毒6 min 后,在无菌操作台内用无菌水洗涤5 次,点播于含有0,100,200,300 mmol/L 甘露醇的MS 培养基中,避光春化3 d 后正常培养4 d,统计发芽率;将野生型与T3 代纯合的过表达株系种子正常点播于MS 培养基上,避光春化3 d 后黑暗培养3 d,取出后置于含有0,100,200,300 mmol/L 甘露醇的MS 培养基中竖直培养7 d 后统计根长;将野生型与T3代纯合的过表达株系种子点播于营养土中,一部分正常培养,另一部分在正常培养4周后停止浇水进行干旱处理,两周后统计表型、测定相关指标,复水4 d后再次统计存活率。
1.2.6 相关指标测定 测定参照的方法:用茚三酮比色法测定游离脯氨酸含量,氮蓝四唑法测定超氧化物歧化酶含量,硫代巴比妥酸比色法测定丙二醛含量。
2 结果分析
2.1 基因GmXTH23的克隆及表达载体构建
以在Phytozome 中查询到的大豆品种Williams 82的基因组序列为参考,设计特异性扩增引物GmXTH23。以大豆品种垦丰16 总cDNA 作为模板,经PCR 扩增得到一条长度为875 bp 左右的序列(图1A)。将目的条带胶回收纯化后连接至pCXSN载体,转化大肠杆菌感受态DH5α并通过菌液PCR鉴定后(图1B)送往测序公司测序,结果与基因GmXTH23参考序列完全匹配。
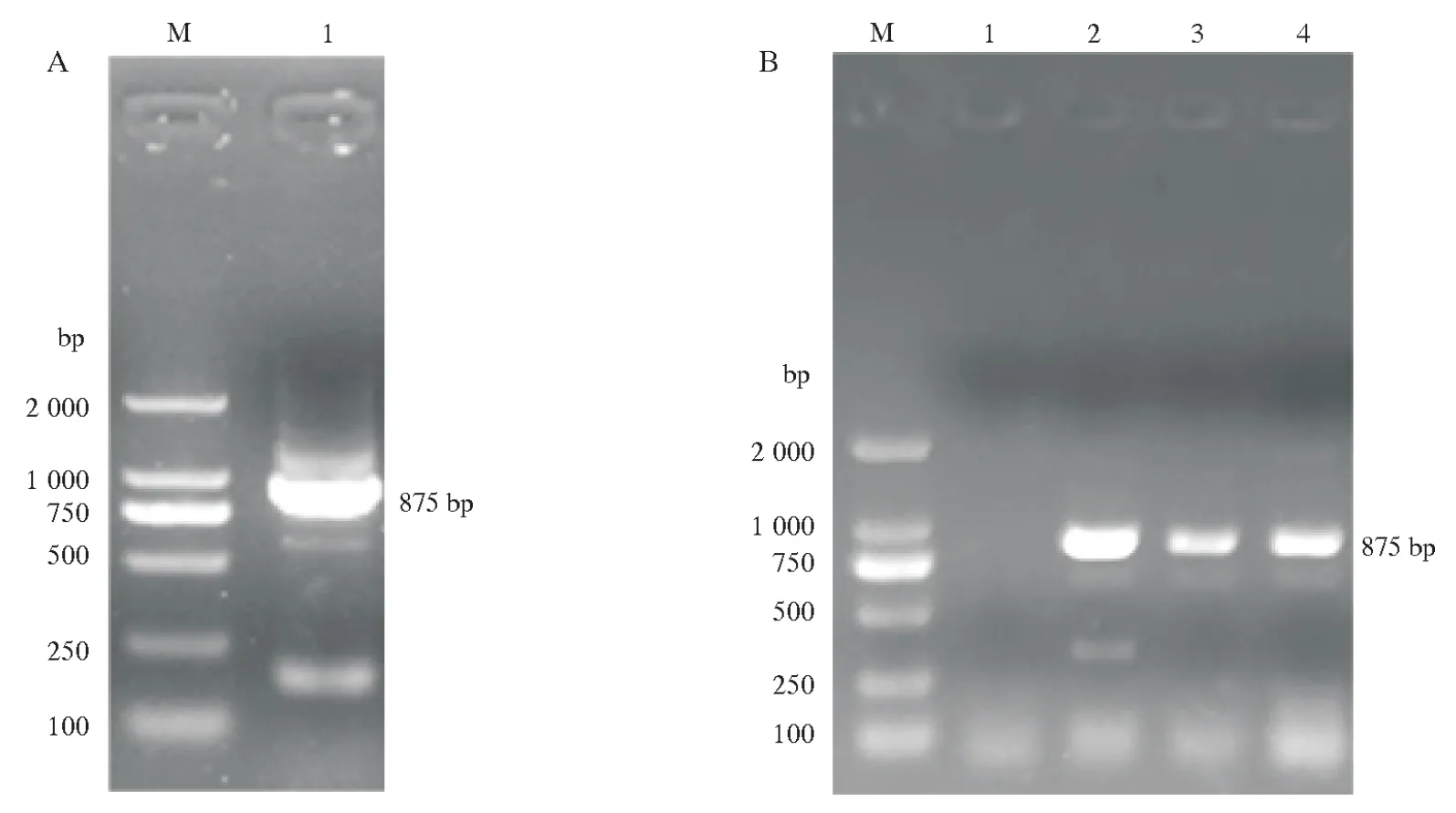
图1 基因GmXTH23的克隆及菌液PCR鉴定Fig.1 Cloning and PCR identification ofGmXTH23gene
2.2 基因GmXTH23不同组织和逆境胁迫下表达模式分析
利用qRT-PCR 对基因GmXTH23在垦丰16不同组织中的表达模式和干旱处理下的响应模式进行分析,结果发现其在大豆的荚、果实、叶片、根部、茎中皆有表达,并且在茎中表达量最高(图2A)。
经5%PEG6000 处理之后,与CK 相比GmXTH23的表达量呈现先升高后下降再逐渐升高的趋势,并在胁迫24 h时达到最高(图2B)。
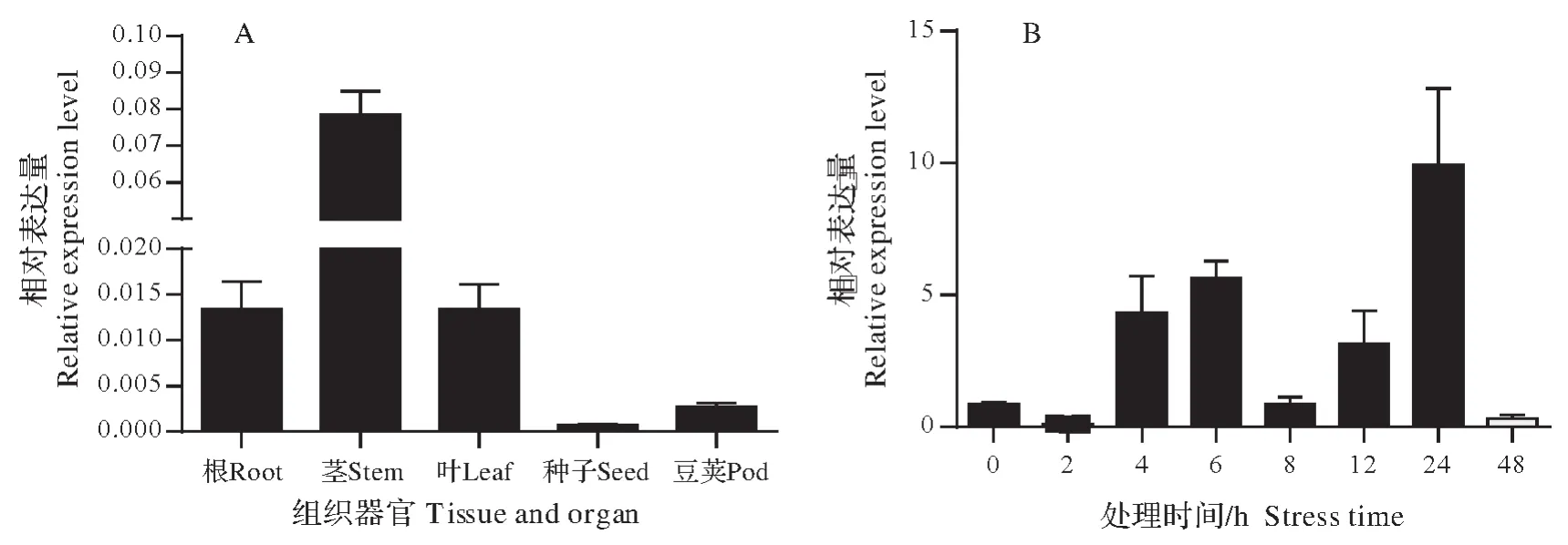
图2 GmXTH23在不同组织间和干旱胁迫后的表达模式分析Fig.2 Analysis ofGmXTH23expression patterns between different tissues and after drought stress
2.3 GmXTH23蛋白的亚细胞定位
为了探究GmXTH23蛋白的定位特征,利用基因枪法将构建好的35S::GmXTH23::YFP融合表达载体转入洋葱表皮细胞内,进行瞬时表达。
通过激光共聚焦显微镜检测发现35S::YFP空载体在整个细胞中皆表达荧光信号,初步确定35S::GmXTH23:YFP荧光信号分布细胞膜上。进一步使用蔗糖溶液进行质壁分离后,再次观察发现GmXTH23-YFP重组蛋白确实定位于细胞膜上(图3)。
2.4 转基因GmXTH23拟南芥抗旱性鉴定
将GmXTH23转基因过表达株系T3代种子与野生型种植于含有0,100,200,300 mmol/L甘露醇的MS培养基中。发现在不同浓度的甘露醇胁迫下,过表达株系的发芽率显著高于野生型(图4)。
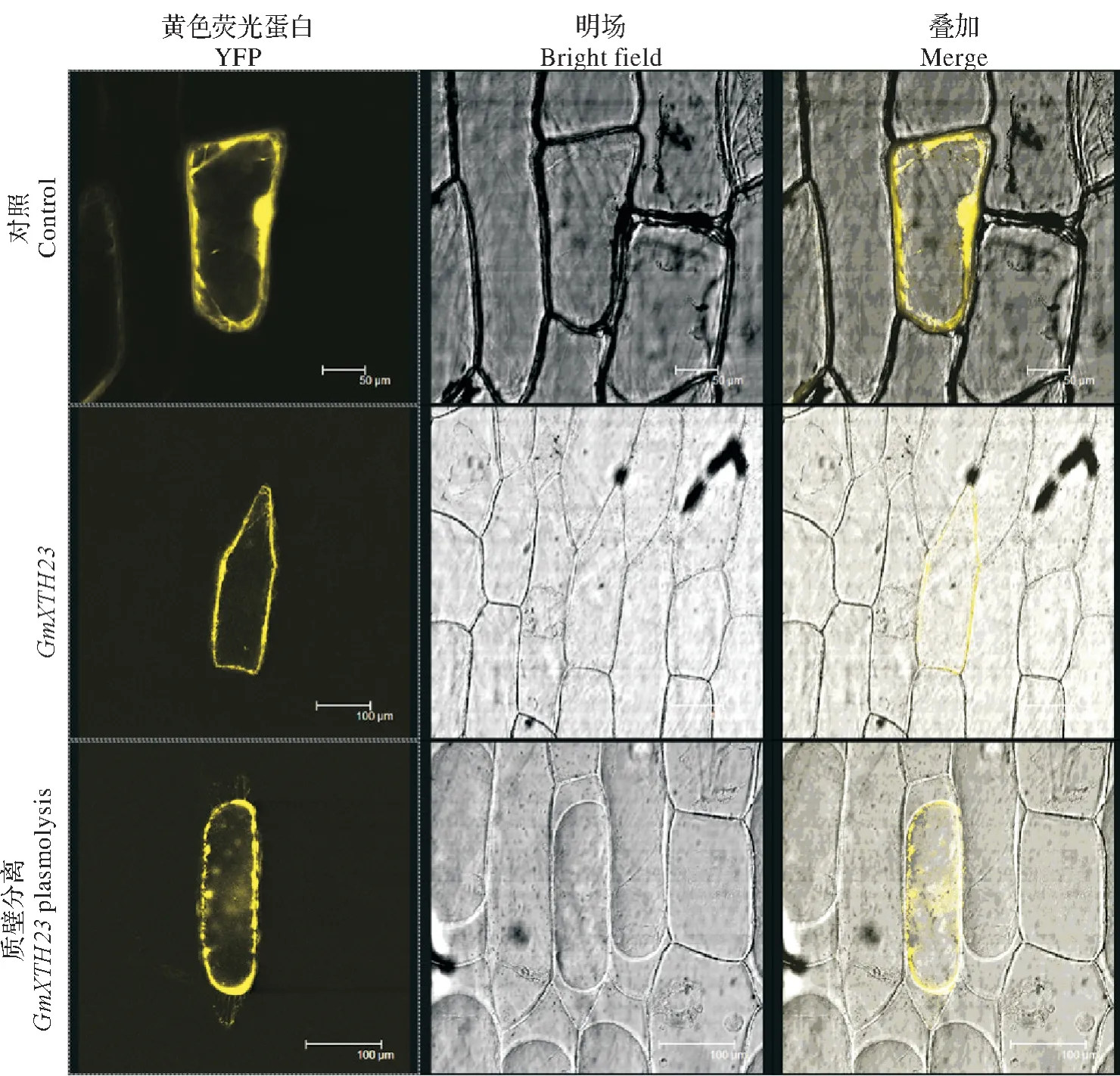
图3 GmXTH23亚细胞定位情况Fig.3 GmXTH23subcellular localization
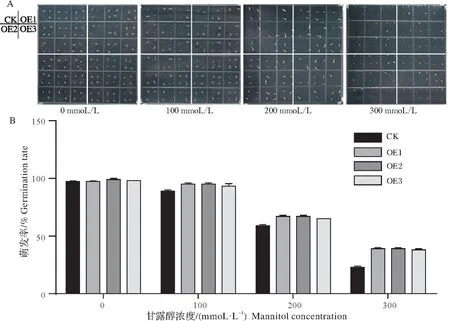
图4 转基因拟南芥干旱胁迫后的萌发情况(A)及萌发率(B)Fig.4 Germination of transgenicarabidopsis thalianaunder drought stress(A)and germination rate(B)
将在正常条件下的MS培养基上培养的T3代GmXTH23转基因拟南芥和野生型拟南芥转移至含有不同浓度甘露醇的MS 培养基中,竖直培养7 d,测量不同株系拟南芥的根长。结果表明在MS 培养基上生长的各株系拟南芥根长基本一致,而在含有不同浓度的甘露醇的MS 培养基中,过表达株系的根长皆显著大于野生型(图5)。

图5 转基因拟南芥干旱胁迫下根生长情况(A)及长度(B)Fig.5 Root growth of transgenicarabidopsis thalianaunder drought stress(A)and length(B)
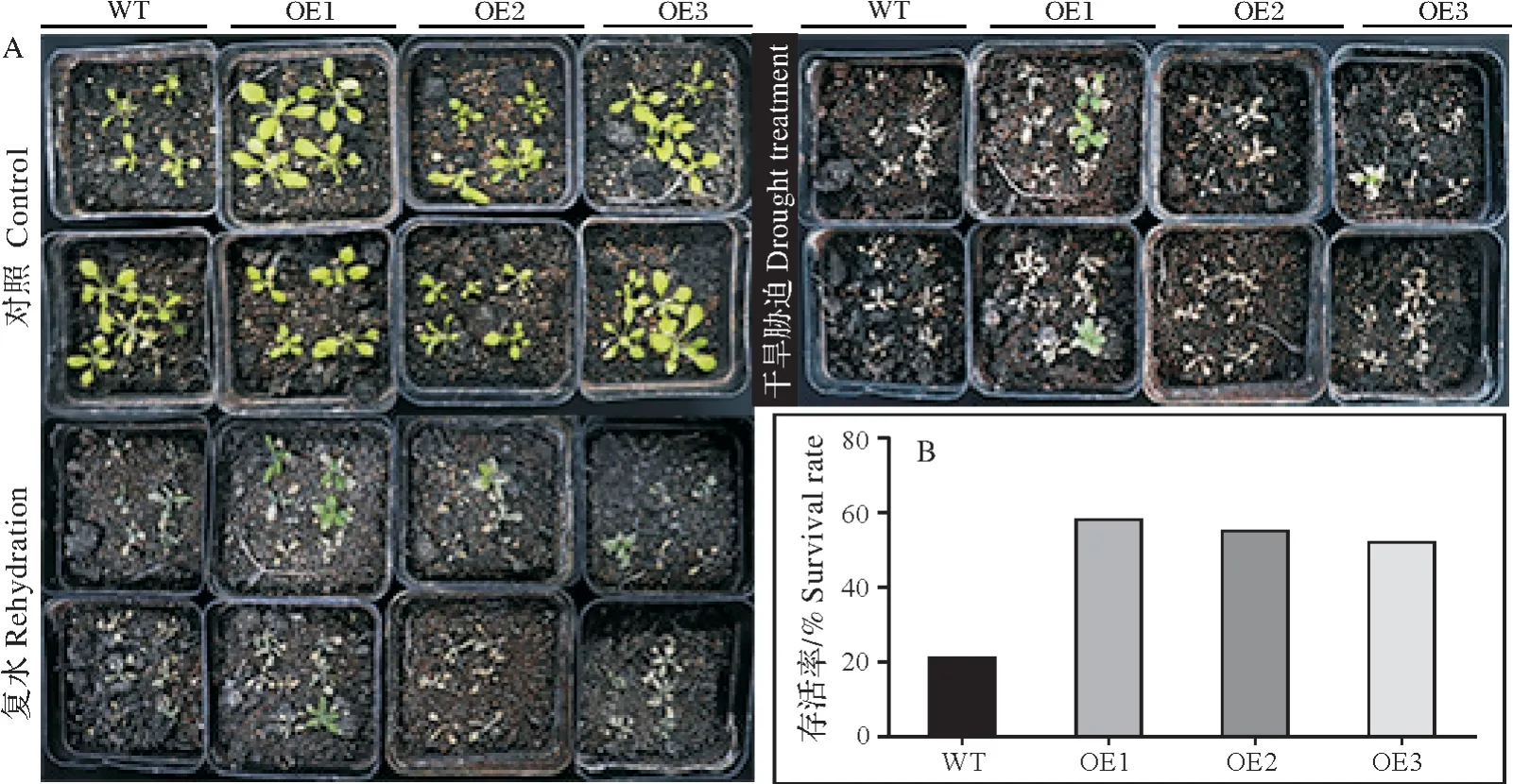
图6 干旱逆境下转基因拟南芥的生长状况(A)和存活率(B)Fig.6 Growth status and survival rate of transgenicarabidopsis thalianaunder drought(A)and stress(B)
将GmXTH23转基因过表达株系和野生型种植在长日照条件下培养四周,没有明显的表型差异。在停止浇水2周后发现四个株系皆出现不同程度的萎蔫,过表达株系OE1、OE2萎蔫程度较轻,OE3程度较重一些,在复水4 d后统计发现58%的过表达植株恢复正常生长,只有18%的野生型植株恢复正常生长(图6)。以上结果表明转GmXTH23基因的过表达株系的抗旱性有所提高。
2.5 相关生理指标的测定
取上述干旱胁迫试验后不同株系的拟南芥叶片,进一步测量其PRO、MDA、SOD含量,发现在经过干旱胁迫后,野生型与过表达株系各项指标皆有差异,过表达株系与野生型株系相比其PRO含量与SOD含量较高(图7A,C),MDA 含量较低(图7B)。结果表明在干旱胁迫下GmXTH23通过提高脯氨酸与超氧化物歧化酶含量、降低植物细胞在胁迫下的损伤来提高植物的抗旱性。

图7 干旱胁迫下转基因拟南芥相关生理指标测定Fig.7 Determination of related physiological indicators of transgenicarabidopsis thalianaunder drought stress
3 讨论
半纤维素是细胞壁构成的主要成分之一,由木葡聚糖构成主链,由植物细胞中的高尔基体合成,囊泡将其转送至细胞膜,与细胞膜结合后分泌到细胞外参与细胞壁的生理代谢[19]。亚细胞定位结果显示,GmXTH23 蛋白定位于细胞膜处,推测其和半纤维素与细胞膜的结合或分泌到细胞外时的生理活动有关。在其他作物中也有类似研究,比如ZmXTH1在分泌途径和细胞壁均有检测到其蛋白,说明其在蛋白的分泌过程中发挥作用[20]。
脯氨酸作为一种较理想的渗透调节物质,不仅可以改善细胞膜的水环境,使膜结构的稳定性增强,还会在植物受到逆境胁迫时在体内大量累积,因此脯氨酸含量可以作为抗旱育种的一项重要生理指标[21]。在干旱处理下,经过测定,相对于野生型,GmXTH23转基因拟南芥含有更高的脯氨酸含量,推测GmXTH23能够通过提高脯氨酸含量来提高植物调节的能力进而提高植物的抗旱性。丙二醛是膜质过氧化作用的产物之一,其含量多少代表了膜质过氧化的严重程度[22];在干旱胁迫下,植物体内会累积较多的活性氧,从而导致细胞遭受膜质过氧化、膜系统被破坏、蛋白质变性等多种细胞毒害。活性氧清除酶促系统中的超氧化物歧化酶(SOD)主要作用就在于它能将超氧阴离子自由基快速歧化为过氧化氢(H2O2)和分子氧;过氧化氢酶(CAT)是清除H2O2的主要酶类,保护细胞避免因累积H2O2而受到损伤,从而控制细胞中的活性氧含量,提高植物抗旱性[23],所以植物体内MDA与SOD的含量也是植物抗旱性的重要生理指标。
本研究初步验证了GmXTH23能提高植物的抗旱性,为大豆抗旱育种提供了理论依据。但是对于基因的生理机制和作用途径尚未清晰,如该基因具体是如何参与细胞壁相关的生理活动、在干旱胁迫下调节机制与途径、在其他胁迫途径中起到何种作用,这些问题对深入探究该基因功能有着启示作用。
致谢:农业部东北大豆生物学与遗传育种重点实验室张宇航和王雪松在试验中给予了指导,谨致谢意!
