沙葱萤叶甲钙结合蛋白基因的鉴定及表达谱分析
2020-11-12周晓榕庞保平单艳敏
李 爽, 李 玲, 周晓榕, 庞保平,*, 单艳敏
(1.内蒙古农业大学草原昆虫研究中心, 呼和浩特 010020; 2.内蒙古自治区草原工作站, 呼和浩特 010020)
Ca2+是一种多功能的第二信使,在生物体的发育过程和对各种环境刺激的反应中起着至关重要的作用(Berridgeetal., 2000)。钙结合蛋白(calcium-binding protein, CaBP)作为Ca2+信号的受体,通过构象改变和活性变化识别并传递特异的Ca2+信号至下游,引起下游特定的反应变化(汪澈等, 2009)。最常见的钙结合蛋白类型是具有1个或多个典型的螺旋-环-螺旋延长因子(EF-hand)基序(helix-loop-helix elongation factor hand motifs)的蛋白,由E螺旋和F螺旋组成,位于钙离子结合环的侧面,该结构类似于手型(Ababou and Zaleska, 2015)。细胞中还有一些蛋白虽然没有EF-hand结构,但依然可以和Ca2+结合,包括与磷酸烯醇丙酮酸羧酶激酶相关的激酶(phosphoenolpyruvate carboxylase kinase-related kinase, PEPRK)、膜联蛋白(annexin)、钙联蛋白(calnexin)、钙网蛋白(calreticulin, CRT)和磷酸酶D(phospholipase D)等。虽然已经对钙结合蛋白的功能开展了研究,但对许多细节和基因组信息仍然缺乏了解(Mohantaetal., 2019)。钙调蛋白(calmodulin, CaM)是真核生物钙结合蛋白EF-hand家族中了解最详细的成员,其参与各种生理活动,包括新陈代谢、免疫反应、细胞凋亡、胞内运动、神经生长和肌肉收缩等(Haeseleeretal., 2002; Tidow and Nissen, 2013)。CaM在昆虫发育变态(Jingetal., 2015)、嗅觉(Bahk and Jones, 2016; Mukundaetal., 2014, 2016)、味觉(Senoetal., 2005)、卵黄生成(Brownetal., 2010; Brubaker-Purkey and Woodruff, 2013; Wangetal., 2018)、抗药性(Guoetal., 2019)以及昆虫与寄主植物相互作用(Hattorietal., 2012; Yeetal., 2017)中可能起着重要作用。钙磷蛋白(calcyphosine, CAPS)是一类在哺乳动物及后生动物中新发现的钙结合蛋白,含有EF-hand基序,与CaM属同一蛋白家族。虽然目前对其确切的功能还不清楚,但可能参与了cAMP和钙-磷脂酰肌醇级联之间的交叉信号传递, 而cAMP和钙-磷脂酰肌醇级联则是控制细胞生长和分化最为重要的途径(鞠川等, 2006)。Lindner等(2019)从脂肪过多症患者体内鉴定出一种类钙磷蛋白(calcyphosine-like, CAPSL),其参与了脂肪的形成。肌钙蛋白(troponin)是调节肌肉收缩的钙感知(Ca2+-sensing)分子开关,是由肌钙蛋白C(troponin C, TnC)、肌钙蛋白T(troponin T, TnT)和肌钙蛋白I(troponin I, TnI)3个亚基构成的三聚复合体,其中TnC为具有EF-hand基序的钙结合亚基,是细胞内钙离子浓度改变诱导肌肉收缩的关键功能组分(Umasuthanetal., 2013)。目前,大部分关于TnC的研究集中于脊椎动物,昆虫TnC的研究很少。Fyrberg等(1994)分离了黑腹果蝇Drosophilamelanogaster的3个TnC亚型;Chen等(2008)研究表明家蚕Bombyxmori的BmTnC在头部、马氏管、体壁和肠道等不同组织中差异表达。CRT是一类内质网中的多功能钙结合蛋白,广泛存在于包括哺乳动物、昆虫、线虫、原生动物及植物等各种生物中,属于(K/H)DEL蛋白家族(Michalaketal., 1992, 1999),在蛋白合成和折叠期间内质网质量控制中起着重要作用(Ramosetal., 2011)。在昆虫中,CRT可能参与嗅觉(Stoltzfusetal., 2003)、免疫反应(Choietal., 2002; Asgari and Schmidt, 2003; Zhangetal., 2006; Wangetal., 2012)及早期发育(Ramosetal., 2011)。滞育是昆虫应对不良环境条件的生活策略,在昆虫生长发育和存活过程中起着重要作用。然而,目前我们对Ca2+信号在昆虫滞育中的调控作用很少了解。新近研究表明,在棉铃虫Helicoverpaarmigera(Lu and Xu, 2010; Zhangetal., 2012)、二斑叶螨Tetranychusurticae(Zhaoetal., 2017)、家蚕(Hsieh and Gu, 2019)和淡色库蚊Culexpipienspallens(Zhangetal., 2019)等滞育过程中许多钙结合蛋白差异表达,意味着其在昆虫滞育调控中可能起着重要作用。
沙葱萤叶甲Galerucadaurica是一种近年来在内蒙古草原上猖獗成灾的新害虫,自2009年在内蒙古锡林郭勒草原突然暴发成灾以来,发生范围不断扩大,危害日趋严重,目前已扩散到锡林郭勒盟、乌兰察布市、呼伦贝尔市、巴彦淖尔市、鄂尔多斯市和阿拉善盟等6个盟市的20多个旗县。该虫发生于荒漠草原和退化草原,取食百合科葱属Allium沙葱A.mongolium、多根葱A.polyrhizum和野韭A.ramosum等植物,使本已脆弱的草原生态环境更加恶化(谭瑶等, 2017)。该虫一年发生1代,春季越冬卵最早于4月上中旬开始孵化,夏季成虫羽化约7~10 d后进入专性滞育,羽化约80~90 d后解除滞育,开始取食、交尾和产卵,以卵滞育越冬(昊翔等, 2015; Zhouetal., 2016; 陈龙等, 2018b)。目前在昆虫纲中以卵冬滞育同时以成虫夏滞育的昆虫未见有过报道,沙葱萤叶甲为研究滞育提供了很好的研究材料。然而,目前对其滞育机理了解得还很少。本实验室前期研究了成虫不同夏滞育阶段体内糖类、蛋白、脂肪含量(陈龙等, 2018b)及海藻糖酶(陈龙等, 2018a)和热激蛋白(陈龙等, 2019)基因表达量的变化。蛋白质组学分析发现,多个钙结合蛋白在沙葱萤叶甲成虫夏滞育不同阶段差异表达(Maetal., 2019)。本研究根据本实验组装的沙葱萤叶甲转录组和蛋白质组数据,应用生物信息学和RT-PCR方法克隆获得沙葱萤叶甲钙结合蛋白基因序列,对其序列特征进行生物信息学分析,并利用qPCR技术检测其在成虫滞育过程中及不同温度下的表达模式,为进一步明确钙结合蛋白在沙葱萤叶甲滞育调控中的作用奠定必要的基础。
1 材料与方法
1.1 供试昆虫
2019年5月于内蒙古锡林郭勒盟镶黄旗采集沙葱萤叶甲幼虫,置于室内自然变温条件下以野韭为食饲养,待羽化为成虫后作为供试虫源。取3日龄成虫雌雄各2头混样作为基因克隆的cDNA模板。qPCR实验包括两类样品:(1)不同温度处理样品:选取羽化后3日龄成虫,分别放置在0, 5, 10, 15, 20, 25, 30及35℃的低温培养箱(LRH-100CB型,上海一恒仪器公司)中处理1 h;(2)不同日龄成虫样品:选取羽化3, 7, 10, 15, 20, 30, 40, 60, 90和110 d的成虫作为供试虫源,其中羽化后3 d的成虫处于滞育前,羽化后7 d开始进入滞育期(7-20 d为滞育初期、30-40 d为滞育中期、60 d为滞育后期),羽化后90 d滞育终止。每个处理3个生物学重复,每个重复雌雄各2头。
1.2 总RNA提取及cDNA的合成
以1.1节收集的虫源为模板,按照RNAiso Plus(TaKaRa,大连)操作说明提取总RNA。提取的总RNA经1.5%琼脂糖凝胶电泳及紫外分光光度计(Nano PhotometerTM-Class,德国)检测总RNA质量、浓度和纯度。分别取1 μg总RNA为模板,按照PrimeScriptTMRT Reagent Kit with gDNA Eraser(TaKaRa,大连)操作说明合成cDNA第1链,放置-20℃保存备用。
1.3 钙结合蛋白基因的克隆
根据本实验室组装的沙葱萤叶甲转录组和蛋白质组数据,筛选关于钙结合蛋白的全部序列,选取4个具有完整开放阅读框(ORF)的钙结合蛋白基因序列,利用Primer Primer 5.0设计引物(表1)。扩增反应体系(25 μL): cDNA模板1 μL, PCR Master Mix 12.5 μL, 上下游引物(0.2 μmol/L)各1 μL, RNase-free Water 9.5 μL。PCR扩增条件: 94℃预变性3 min; 94℃变性30 s, 58℃退火30 s, 72℃延伸 30 s, 30个循环; 72℃延伸10 min。扩增产物用1.5%的琼脂糖电泳检测后,胶回收目的片段。将回收的目的片段与pMD®19-T连接,转入大肠杆菌EscherichiacoliDH5α感受态细胞,经蓝白斑筛选, 挑选白色菌落进行PCR验证,将验证过的菌液继续过夜培养过夜后,送至上海生工公司进行测序。

表1 引物信息
1.4 序列分析及系统发育树构建
利用NCBI数据库中ORF Finder对沙葱萤叶甲钙结合蛋白基因的开放性阅读框进行预测(https:∥www.ncbi.nlm.nih.gov/orffinder/);利用DNAMAN 6.0软件进行分子量及理论等电点的预测;利用SingalP 5.0软件进行信号肽预测(http:∥www.cbs.dtu.dk/services/SignalP/);利用TMHMM Server 2.0(http:∥www.cbs.dtu.dk/services/TMHMM/)进行跨膜区的预测;利用Scan Prosite(https:∥prosite.expasy.org/)对蛋白结构域预测。利用MEGA 6.0软件的邻接法(重复运行1 000次)构建系统发育树。
1.5 钙结合蛋白基因表达谱qPCR分析
qPCR反应体系(20 μL): 模板cDNA 2 μL, 上下游引物(10 μmol/L)各0.4 μL, GoTaq®qPCR Master Mix(2×)10 μL(Promega,美国), Nuclease-Free Water 7.2 μL。选用沙葱萤叶甲SDHA(GenBank登录号: KU240575)作为内参基因(Tanetal., 2017),每个处理3次生物学重复及3次技术重复。使用qPCR仪(FTC-3000,Funglyn Biotech,加拿大)进行测定。反应程序: 95℃预变性10 min;95℃变性15 s, 60℃退火1 min, 40个循环; 95℃变性15 s, 60℃退火15 s, 95℃延伸15 s。
1.6 数据分析
采用2-ΔΔCt法(Livak and Schmittgen, 2001)测定沙葱萤叶甲钙结合蛋白的相对表达量,不同日龄成虫及不同温度下钙结合蛋白的相对表达量差异显著性分析采用单因素方差分析(one-way ANOVA)和Duncan氏新复极差法。运用SPSS 22.0统计分析软件对数据进行分析,数据以平均值±标准误表示。
2 结果
2.1 沙葱萤叶甲钙结合蛋白基因的鉴定及序列特征
通过对沙葱萤叶甲成虫转录组数据的BlastX比对分析,经RT-PCR克隆、测序验证及生物信息学分析获得4个具有完整开放阅读框(ORF)的钙结合蛋白基因序列,依次命名为GdCaM,GdCAPSL,GdTnCl和GdCRT(GenBank登录号:MN695412-MN695415)。ORF全长分别为450, 648, 516和1 209 bp,分别编码149, 215, 171和402个氨基酸残基,预测蛋白质分子量分别为16.81, 24.34, 19.67和46.43 kD,等电点pI分别为3.84, 4.57, 3.95和4.17。GdCaM, GdCAPSL和GdTnCl无信号肽,分别与玉米根萤叶甲DiabroticavirgiferavirgiferaCaM, CAPSL和TnCl的氨基酸序列一致性最高,分别为100%,74.0%和88.2%;GdCRT与马铃薯甲虫LeptinotarsadecemlineataCRT的氨基酸序列一致性最高(92.5%)。根据EF-hand基序的共有序列D-x-DNE-x-DNS-GP-x-ILVFM-DSE-x-x-DE(x代表任意氨基酸),运用Scan Prosite在线软件分析显示(图1):GdCaM有4个典型的结构和功能性EF-hand基序;GdCAPSL有3个EF-hand基序,其中EF-hand 4基序缺失[EF-hand基序氨基酸被替代,其中第1位的甘氨酸(Gly)替代了天冬氨酸(Asp),第3位的苏氨酸(Thr)替代了天冬氨酸/天冬酰胺(Asn)/谷氨酸(Glu)];GdTnCl只有2个EF-hand基序,其中EF-hand 1和EF-hand 3基序均缺失[EF-hand 1基序第5位的赖氨酸(Lys)替代了天冬氨酸/天冬酰胺(Asn)/丝氨酸(Ser),第6位的谷氨酰胺(Glu)替代了甘氨酸(Gly)/脯氨酸(Pro),第12位的蛋氨酸(Met)替代了谷氨酸(Glu)/天冬氨酸(Asp);EF-hand 3基序第5位的亮氨酸(Leu)替代了天冬氨酸/天冬酰胺(Asn)/丝氨酸(Ser),第12位的亮氨酸替代了谷氨酸(Glu)/天冬氨酸(Asp)]。GdCRT不具有EF-hand基序,但GdCRT具有2条典型的钙网蛋白(CRT)家族标签序列(K97HEQNIDCGGGYV KVF112和L129MFGPDiCG137)、三重复序列(I207kDPEAKKPEDWD219, I224PDPDDTKPEDWD236和I241PDPDATKPDDWD253)及内质网滞留信号序列HDEL(图2)。
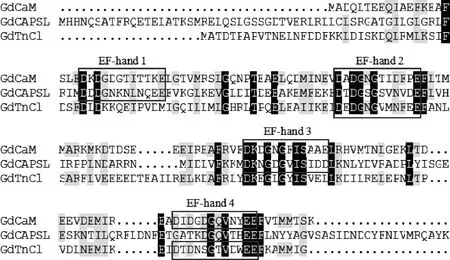
图1 沙葱萤叶甲EF-hand钙结合蛋白序列分析
2.2 沙葱萤叶甲及其他鞘翅目昆虫钙结合蛋白的系统发育关系
将沙葱萤叶甲钙结合蛋白氨基酸序列与NCBI数据库中已知的其他鞘翅目钙结合蛋白氨基酸序列构建系统树,结果图3表明:具有EF-hand基序的GdCaM, GdCAPSL和GdTnCl分别与其他鞘翅目昆虫的CaM, CAPSL和TnCl各构成一个分支,其中GdCaM置信度均为100%,在进化过程中具有高度保守的进化特征;不同昆虫TnCl间保守性较高,置信度介于47%~99%之间,其中GdTnCl与玉米根萤叶甲的TnCl亲缘关系最近,最先聚在一起。这3类具有EF-hand基序的钙结合蛋白亲缘关系较近,共同聚为一个大分支,最后与CRT聚在一起。GdCRT与其他鞘翅目昆虫的CRT单独聚为一个分支,但不同昆虫CRT间的亲缘关系较远,置信度较低。

图3 邻接法构建的基于氨基酸序列的沙葱萤叶甲及其他鞘翅目昆虫钙结合蛋白系统发育树(1 000次重复)
2.3 沙葱萤叶甲钙结合蛋白基因在不同日龄成虫中的表达分析
qPCR结果(图4)表明,GdCaM,GdCAPSL,GdTnCl和GdCRT在不同日龄成虫中的表达量存在显著差异(P<0.05),且表达模式不同。GdCaM在成虫滞育前(羽化后3 d)表达量最高,开始进入夏滞育后(羽化后7 d)表达量降低,在滞育期间(羽化后7-60 d)变化较小,而解除滞育后(羽化后90 d)表达量又显著下调(图4: A)。GdCAPSL在成虫羽化后表达量逐渐下降, 在滞育初期(羽化后10 d)维持在最低水平,进入滞育中后期(羽化后40 d和60 d)开始回升,滞育解除后又突然下调至最低水平,而羽化后110 d急剧上升至最高水平(图4: B)。GdTnCl随成虫发育上调表达,在滞育初期(羽化后15-20 d)达最高水平,进入滞育中后期(羽化后30-60 d)急剧下降至最低水平,滞育解除后再次上调,但在羽化后110 d突然下调至最低水平(图4: C)。GdCRT在进入滞育后表达量开始逐渐下调,在滞育维持期(羽化后15-60 d)维持在低水平,滞育解除后又开始上升(图4: D)。
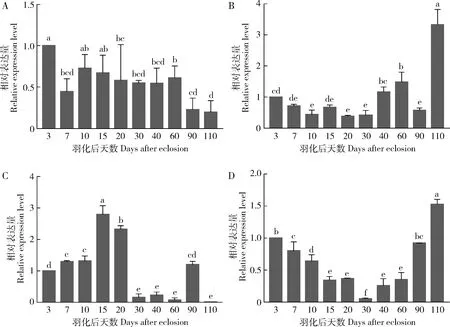
图4 沙葱萤叶甲不同日龄成虫体内钙结合蛋白基因的相对表达量
2.4 沙葱萤叶甲钙结合蛋白在不同温度胁迫下的表达分析
从图5可知,温度对除GdCRT外的其他3种钙结合蛋白基因(GdCaM,GdCAPSL和GdTnCl)表达量均有显著影响(P<0.05)。当温度高于20℃,随着温度的逐渐上升,GdCaM的表达量逐渐上升;当温度低于20℃时,随着温度的逐渐降低,GdCaM的表达量上调,在5℃达到最高值,但0℃时又突然下降到最低值(图5: A)。GdCAPSL的表达量随着温度的升高而呈现上升的趋势,25℃时达到最高,然后下降(图5: B)。GdTnCl在0℃时表达量最高,随着温度的升高,表达量呈现整体降低的趋势(图5: C)。
3 讨论
根据沙葱萤叶甲转录组和蛋白质组数据,本研究首次鉴定和克隆得到4条具有完成ORF的钙结合蛋白基因cDNA序列,其中3条编码的蛋白属于具有典型EF-hand结构域的钙结合蛋白家族,另一条编码的蛋白为不含EF-hand结构域但具有钙结合能力的钙网蛋白(CRT)。氨基酸序列比对分析表明,沙葱萤叶甲GdCaM序列与其他鞘翅目昆虫CaM的完全相同,均具有4个典型的EF-hand基序,进一步说明CaM具有高度保守的进化特征(李庆伟等, 2017)。我们从沙葱萤叶甲转录组数据库BLAST获得一条类似钙磷蛋白(calcyphosine-like, CAPSL)的序列,其与同属萤叶甲亚科的玉米根萤叶甲CAPSL一致性最高(74%),命名为GdCAPSL。结构域分析表明,GdCAPSL具有3个典型的EF-hand基序,而CAPS与CaM一样具有4个EF-hand基序(鞠川等, 2006)。TnC也是具有4个典型的EF-hand基序的钙结合蛋白,但由于一些TnC中某些基序上的钙离子结合所必需的氨基酸残基发生突变,使得其失去2个甚至更多的钙离子结合位点(Qiuetal., 2003)。本研究获得的沙葱萤叶甲TnC失去2个钙离子结合位点,只剩2个EF-hand基序,且与玉米根萤叶甲TnCl的氨基酸序列一致性最高(88.2%),因而命名为GdTnCl。CRT是不具有EF-hand结构域的钙结合蛋白,但至少具有3个典型的功能结构域:(1)N端结构域:具有一段信号肽、2条保守的钙网蛋白家族标签序列KHEQNIDCGGGY和IMFGPDICG;(2)P端结构域:富含脯氨酸,并有1个高亲和性、低容量的钙离子结合位点和2组三重复序列;(3)C端结构域:拥有一个高容量、低亲和性的钙离子结合位点和典型的内质网滞留信号肽(K/H)DEL(Michalaketal., 1992, 1999)。生物信息学分析表明,本研究克隆获得的一段ORF编码的蛋白序列具有上述特征,而且与马铃薯甲虫CRT氨基酸序列一致性高达92.5%,因此将该序列命名为GdCRT。
CaM是一种重要的调节蛋白,能结合第二信使Ca2+和多种靶蛋白,调节多种细胞过程和发育。GdCaM在沙葱萤叶甲成虫滞育前(羽化后3 d)的表达量最高,随后在进入滞育后(羽化后7 d)表达量显著下降(图4: A),这一规律与棉铃虫幼虫滞育过程CaM水平降低(Lu and Xu, 2010; Zhangetal., 2012)相一致;淡色库蚊滞育的各阶段中CaM表达也呈现显著变化,表明CaM可能在滞育过程中起重要的调控作用(Zhangetal., 2019)。但在解除滞育后,GdCaM表达量进一步下降的原因尚不清楚。Teets等(2013)发现钙信号介导昆虫冷感应,当25℃下降到0℃,细胞内钙离子浓度增加40%,并使得钙调蛋白依赖性蛋白激酶Ⅱ(CaMKII)活性升高。与本研究中发现短时高温和低温胁迫均导致GdCaM表达量上调,而0℃时GdCaM的表达量低于5℃时的(图5: A),可能是过低的温度使得GdCaM表达受到抑制。
CAPSL也属于钙结合蛋白EF-hand家族,可能与CaM存在相互作用(Klimmecketal., 2008)。CAPSL在沙葱萤叶甲成虫滞育中后期(羽化后40-60 d)表达量显著上调(图4: B)。Zhao等(2017)也发现,与非滞育对照相比,CAPSL在二斑叶螨滞育第3和第13天表达量显著上调。说明GdCAPSL可能与沙葱萤叶甲滞育调控有关。
TnC通过结合钙离子发挥其生物学功能,TnC随着细胞内Ca2+浓度的升高,引起肌动蛋白与肌球蛋白发生相互作用,产生肌肉收缩或扩张(Filatovetal., 1999),肌肉结构的稳定性在寒冷生存过程中起着至关重要的作用(Zhaoetal., 2017)。本研究中发现,GdTnCl的表达量随着温度的降低而上升(图5: C),表明GdTnCl可能通过参与调节肌肉结构的稳定性,进而提升昆虫在寒冷条件下的适应能力。GdTnCl在沙葱萤叶甲成虫滞育初期(羽化后7-20 d)表达量上调,在滞育中后期(羽化后30-60 d)下调表达(图4: C)。尖音库蚊Culexpipiens中发现肌动蛋白(actin)在滞育初期表达量上调,滞育后期又回到低水平(Kimetal., 2006)。GdTnCl可能通过对肌动蛋白的作用间接调控细胞骨架系统,引起细胞骨架系统重排,通过结构组分改变对沙葱萤叶甲夏滞育做出反应。
GdCRT在沙葱萤叶甲成虫滞育期间(羽化后7-60 d)的表达量显著低于滞育前(羽化后3 d)和解除滞育后(羽化后90 d)(图4: D),CRT可能通过表达量的下调来调控神经递质的释放,进而诱导昆虫进入滞育。在棉铃虫滞育期幼虫滞育诱导过程中,同样发现CRT表达量水平下降(Lu and Xu, 2010)。另外,沙葱萤叶甲成虫羽化110 d时GdCAPSL和GdCRT的表达量达到最高值,而GdTnCl的表达量降为最低值,此时雌雄成虫已经进行交尾受孕,我们推测对应的3种CaBP可能在生殖过程中也发挥着潜在作用。
本研究首次成功克隆了4个沙葱萤叶甲钙结合蛋白基因,并进行了分子特性及在成虫发育过程中和不同温度下的表达分析,为进一步明确钙结合蛋白在沙葱萤叶甲发育和成虫滞育中的作用奠定了基础。
